Population Viability Analysis of Riverine Fishes
Henriëtte I. Jager
Oak Ridge National Laboratory
Oak Ridge, TN 37831-6036
Ken Lepla, James Chandler, and Phil Bates
Idaho Power Company
Boise, ID 83702
Webb Van Winkle
Oak Ridge, TN 37830
Abstract
Many utilities face conflicts between two goals: cost-efficient hydropower generation and protecting riverine fishes. Research to develop ecological simulation tools that can evaluate alternative mitigation strategies in terms of their benefits to fish populations is vital to informed decision-making. In this paper, we describe our approach to population viability analysis of riverine fishes in general and Snake River white sturgeon in particular. We are finding that the individual-based modeling approach used in previous in-stream flow applications is well suited to addressing questions about the viability of species of concern for several reasons. Chief among these are: (1) the ability to represent the effects of individual variation in life history characteristics on predicted population viability; (2) the flexibility needed to quantify the ecological benefits of alternative flow management options by representing spatial and temporal variation in flow and temperature; and (3) the flexibility needed to quantify the ecological benefits of non-flow related manipulations (i.e., passage, screening, and hatchery supplementation).
Introduction
Understanding the relationships between stream habitat and threatened or endangered fish species requires a flexible modeling approach. We are experiencing a shift in attention from abundant, augmented populations of game species to native fishes that have declined in abundance and may now be at risk of extinction. During this transition, we discovered three main differences between previous population models and those suitable for population viability analysis (PVA). First, to accommodate a longer time horizon, it was appropriate to reduce the spatial and temporal detail of our river models. Population viability analysis seeks to predict the chance that a population will persist over a long time horizon. We replaced the fine-scale simulation approach of our in-stream-flow models with a simplified way of representing spatial variation and temporal dynamics. Second, new variables are relevant at long time scales that are not relevant at shorter time scales. For example, when possible, PVAs consider the effect that reduced genetic variation may have on the extinction risk of small, isolated populations. Our individual-based approach is pre-adapted to simulate genetic diversity and feedback between genetics and population viability. Third, the dynamics of small populations may differ from those of larger populations. Several authors of this paper have a history of research focused on compensatory density-dependent mechanisms in natural populations. When populations fall below a critical density, they can experience depensatory population growth. These "Allee" effects are best illustrated by disease eradication programs. By inoculating a significant proportion of potential hosts, these programs seek to cause the extinction of the disease organism. When the density of disease organisms is low enough, a positive feedback between density reduction and the rate of population decline leads to eradication. A potential depensatory mechanism in sturgeons and other broadcast spawners is the decline in egg fertilization rates as spawning aggregations become smaller.
PVA models typically represent environmental variation as random fluctuations in demographic parameters. This approach does not adequately address trends in environmental change or the relationship between policy and population persistence. Therefore, we decided to preserve our focus on linking the flow environment to fish biology. In this paper, we will describe the importance of this linkage in PVA models of riverine fishes.
The goals of this paper are (1) to describe the current status of white sturgeon populations in the Snake River; (2) to outline our approach to environmental PVA; and (3) to highlight the results of several preliminary simulation experiments. These experiments improved our understanding of the role played by the following factors on population viability: fragmentation, migration, habitat augmentation, demographic and genetic variation.
Snake River White Sturgeon
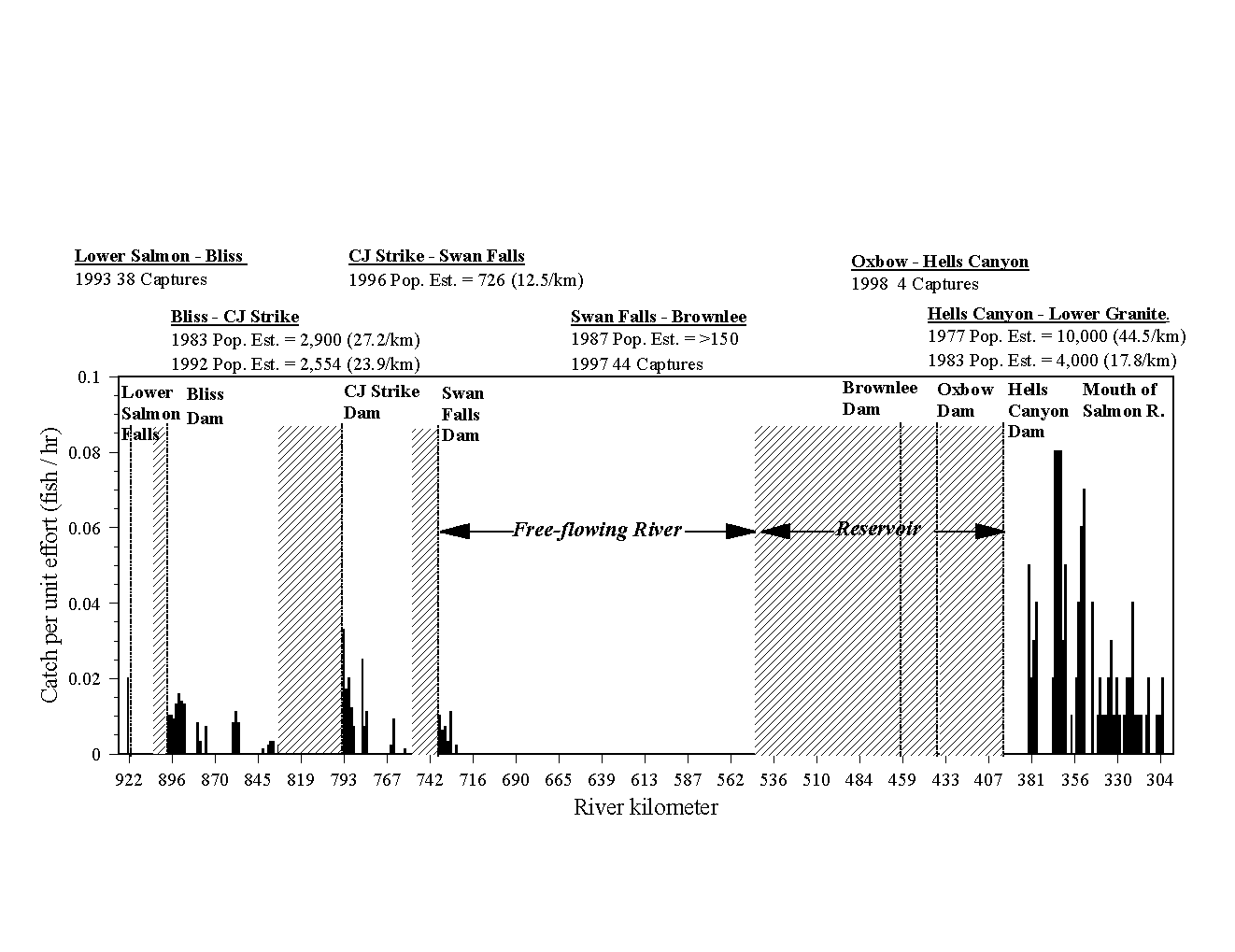
The white sturgeon (Acipenser transmontanus) is a large, long-lived and late-maturing fish species that historically migrated between the estuaries along the Pacific coast and large river systems. From north to south, these include the Fraser, Columbia, San Joaquin and Sacramento rivers. The longest of these is the Columbia River, which drains about 260,000 sq km. The Snake River, the largest tributary to the Columbia, runs from Wyoming to the Washington coast. The Snake River offers a contrast in population status between river segments with healthy, reproducing populations and others with very few individuals and no detectable recruitment (Figure 1). The river segments below Hells Canyon and Bliss dams have sizable populations and show signs of recent reproduction. Reaches with small populations and little or no detectable reproduction occupy the middle sections of the river between Hells Canyon and Swan Falls dams and upstream of Bliss Dam. Several hydroelectric projects along the Snake River have been involved in re-licensing since 1990. To anticipate and address issues raised as part of re-licensing we are conducting PVA analysis to assess the viability of white sturgeon populations in the Snake River.
Figure 1. The current status of white sturgeon populations in the Snake River.
Linking Extinction Risk to Hydrologic Conditions
Because river flow has important effects on fish populations in the river environment, representing linkage between annual hydrologic conditions and population demographic parameters should be an important focus of riverine PVAs. The PVA asks how long the population is likely to persist into the future with a continuation of historical hydrologic patterns. We plan to characterize typical sequences of hydrologic years by dividing the historical record of annual river flow into dry, normal, and wet years. Thus, we will simulate future environmental conditions in the Snake River by simulating stochastic sequences of hydrologic year types according to a Markov model (Table 1).
Table 1.
Markov transition probabilities used to simulate future hydrology
in the Snake River, Idaho.
|
|
To hydrologic-year type |
||
|
From type |
Dry |
Normal |
Wet |
|
Dry |
0.68 |
0.18 |
0.14 |
|
Normal |
0.43 |
0.38 |
0.19 |
|
Wet |
0.28 |
0.33 |
0.39 |
Our next challenge will be to quantify how demographic parameters differ during different hydrologic years (Table 2). We explicitly represent two mechanistic sources of mortality on white sturgeon that depend on temperature and river flow. First, the survival of eggs and larvae through spring depends on water temperatures remaining below critical levels until they develop into young-of-year fish. We will estimate the survival to age one in wet vs. dry years based on a relationship between survival and temperature for daily temperature series typical of each hydrologic year class during incubation. Second, survival of all ages through summer conditions in the reservoir environment depends on maintaining sub-lethal temperature and adequate dissolved oxygen. The amount of suitable habitat available during the hottest part of summer in a given river segment and hydrologic year type will influence susceptibility to density-dependent mortality.
In addition to simulating the effects of hydrology on survival, we are linking hydrology with reproduction and migration (Table 2). For projected years of each hydrologic type, the PVA model will apply weighted usable area for spawning averaged over historical years of the appropriate hydrologic type. We will represent the effects of hydrology on migration and risks associated with migration by increasing downstream migration with increasing flow. This flow-mediated subsidy of downstream populations will be amplified by increased survival of migrants during wet years when a higher proportion of flow is spilled rather than routed through turbines.
Table 2.
Simulated links between annual hydrology and white sturgeon survival and reproduction.
|
Life stage and process |
Description of conceptual PVA model |
|
Reproduction |
Wetter years provide better hydrologic conditions for spawning and dispersal of fertilized eggs. |
|
Survival of post-hatch larvae |
Chances of settling to the bottom before washing downstream out of the reach decrease with turnover rates, which depend on the hydrologic year type. |
|
Survival of juvenile and adult sturgeon |
Risk of mortality caused by episodes of summer anoxic conditions depends on the proportion of reservoir volume with suitable temperature and DO (a function of hydrologic year type) and the amount of suitable free-flowing riverine habitat to provide refuge. |
|
Migration |
Chances of survival during migration depend on fish size and the risk of entrainment, which depends on the hydrologic year type. |
In simulation experiments, we showed that one-way migration can adversely affect persistence of upstream sub-populations. We simulated entrainment mortality as a function of increasing fish size. In these simulations with low levels of downstream migration, entrainment mortality had only a small effect on overall persistence in both wet and dry years (Figure 2). As one would expect, entrainment mortality only influenced the likelihood of persistence of downstream sub-populations.
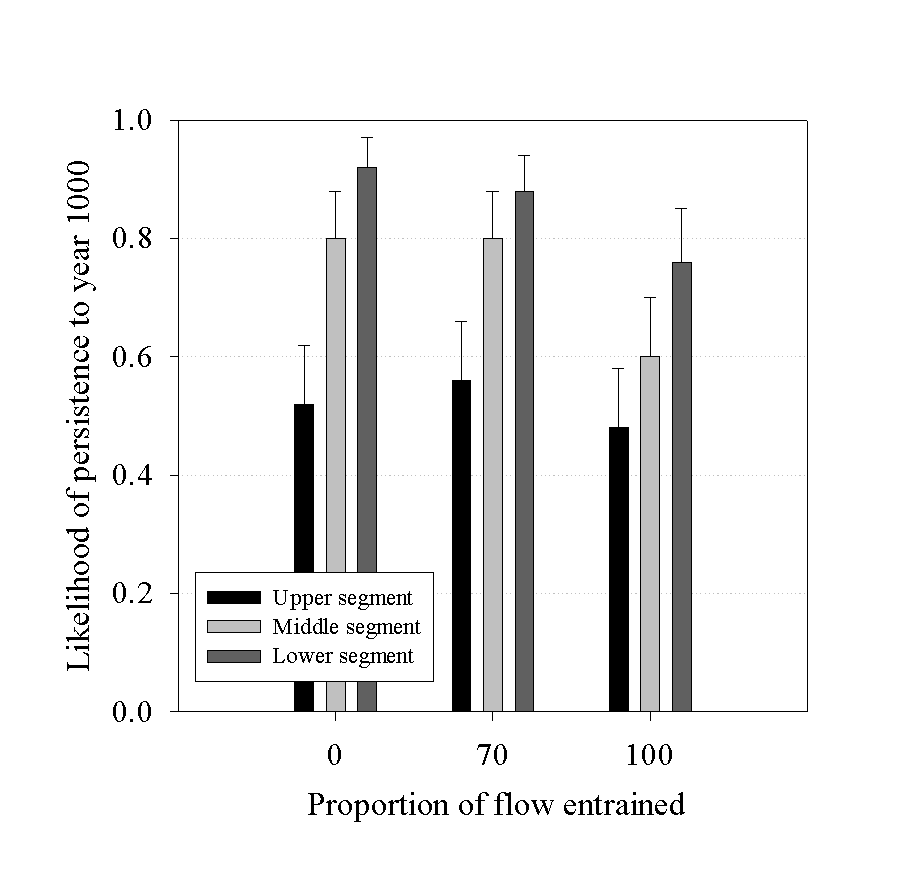
Figure 2. Simulated likelihood of persistence of white sturgeon in three linked river
segments. Entrainment levels correspond to a low (100%) and a high (70%) flow year.
Viability Analysis of Riverine Metapopulations
The Snake River system (Figure 1) consists of a number of linked river segments separated by dams. Strictly speaking, this is not a metapopulation because there is no upstream migration. With either translocation ("assisted fish movements") or provision for upstream passage, these Snake River white sturgeon populations would form a metapopulation. This would open the door for potential management and conservation as a metapopulation, rather than as a series of isolated populations, each with adequate habitat diversity to support all lifestages through a reasonable range of future environmental conditions. The metapopulation approach recognizes that many riverine species evolved to follow a "conveyor belt" life history that combined upstream movement to reproduce with downstream movement as juveniles. Incorporating movements into the developmental cycle ensured that each life stage would encounter appropriate habitat along the longitudinal spectrum of available habitats.
To understand the effects of population isolation (fragmentation) and habitat availability, we conducted simulation experiments using our 1st year version of the white sturgeon PVA model. The first simulation experiments began with a fixed length of river partitioned into equal-length segments by dams. The first simulation had one dam, the second, two dams, etc. Our confidence in the parameter values used is not high enough to advocate using these results to estimate the minimum length of river needed to sustain a white sturgeon population. However, we do feel confident that the pattern illustrated by Figure 3 describes the qualitative effect of habitat fragmentation on the viability of sub-populations. We determined that the likelihood of persistence declined as we divided the river into a larger number of small isolated sub-populations.
In a second experiment, we considered the added effect of habitat loss. In addition to isolating populations, constructing dams converts free-flowing-river habitat to reservoir habitat. We simulated two possible adverse effects on white sturgeon populations that are associated with reservoir habitat. First, reservoir habitat is less-likely than riverine habitat to be suitable for spawning. Second, the risk of exposure to anoxic summer conditions is higher in storage reservoirs, particularly when aggravated by high nutrient loading from agricultural runoff. Our simulations indicate that the simulated length of river habitat needed to maintain a viable population increased (Figure 3). However, for low and intermediate numbers of dams, loss of habitat had no effect on the likelihood of persistence.
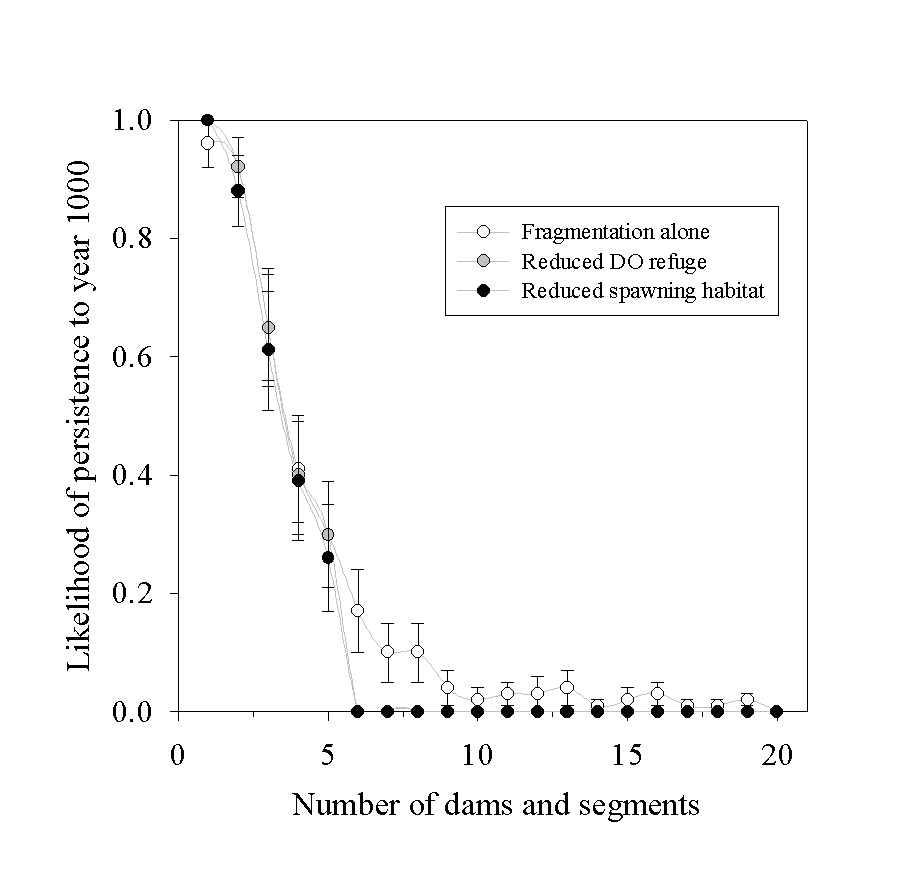
Figure 3. Fragmentation of the river habitat decreased the chances for population persistence. Associated habitat reduction (i.e. due to less refuge from low DO or reduced spawning habitat) had no effect in longer river reaches, but led to extinction in simulations of highly fragmented rivers.
A third simulation experiment evaluated the effects of migration between three linked river segments on the likelihood of persistence. When we simulated migration among the three sub-populations, we found that the likelihood of persistence increased as long as levels of upstream and downstream migration were low and approximately equal. These results suggest that relocating individuals or providing passage from downstream to upstream populations can rescue upstream populations from extinction.
Individual Variation
Individual white sturgeon vary a great deal in their life history characteristics. The age at first maturity of females can vary from 11 to 34 years! Clearly, it would be a challenge for a lumped-parameter population model to simulate population dynamics accurately by assigning the same fecundity to all individuals in an age or size category when this much variation is present. Past studies of individual variation tended to report an increase in population size and viability with increased variability among individuals. For example, individual variation in acquiring food resources allowed persistence at much smaller population sizes than simulations of species with a more egalitarian social system (i.e., no individual variation or territoriality). Likewise, in size-structured fish communities, individual variation in growth resulted in higher survival. In contrast, individual differences in ages at maturity did not necessarily enhance the viability of simulated white sturgeon populations. Rather, the outcome depended on whether the value assigned in simulations of constant age at maturity was optimal in terms of current environmental conditions. While neglecting individual variation can have a significant effect on predicted future viability, we are not yet in a position to generalize about the direction of the effect. In contrast, neglecting individual variation with a genetic basis will always cause predictions of population viability to be low in a changing environment.
Genetic Risks
In addition to demographic concerns, genetic considerations should be addressed by PVA. Usually, this is because small populations are subject to Allee effects caused by inbreeding depression and loss of genetic variability. Another important role of genetic models is to provide information on the costs and benefits of supplementing fish populations with hatchery reared stock.
Because changes to the river environment associated with dam building and hydropower generation can change the selective regime acting on fish species, the role of genetic change (i.e., evolution) in life history traits can be relevant. When there is sufficient genetic diversity, allowing life histories to evolve decreases the projected likelihood of population extinction. Changes in climate associated with fossil fuel burning provide the best-studied example of a shifting selective regime. Theory suggests that, with enough genetic and phenotypic variation and sufficiently slow change, environmental changes may be compensated for by evolution in seasonal cues or thermal tolerances. Like businesses, fishes are better able to adapt to changes when they are predictable.
Our sturgeon PVA model can track both neutral genetic variation and that associated with specified traits that influence fitness of individual fish in a modified selective regime. So far, we have used the model to quantify the loss of genetic diversity with increased fragmentation. In a more theoretical application, the model quantified the role of demographic and genetic variation in life history traits in a modified riverine environment. In future, we may consider a third potential application. There is great interest in the hatchery supplementation. The model described here would be capable of quantifying the genetic costs, as well as the demographic benefits, of transplanting fish bred from more-distant populations. For example, those with ocean access below Bonneville Dam or even the population with threatened status in the Kootenai River.
Conclusions
Individual-based population models provide a good foundation for simulating the future effects of alternative management scenarios. This is because IBMs have the flexibility to represent individual differences in age, size, spatial location, and other relevant attributes. Future simulations can be designed to quantify the mitigative value of hatchery supplementation, the influence of fishing regulations on population viability, and the value of habitat improvements that target specific life stages of white sturgeon. We envision that this approach is suitable for other riverine fish species of concern with similar ties to the river environment and facing similar stresses and mitigation alternatives.
Acknowledgements
We appreciate helpful review comments provided by Mark Bevelhimer. We thank the white sturgeon research community in the Columbia River basin for participating in workshops. This research is sponsored through a tailored collaboration between the Electric Power Research Institute and Idaho Power Co. under EPRI contract WO9117-01 (DOE No. ERD-95-1332) with Lockheed Martin Energy Research Corp. This is Environmental Sciences Division Publication No. 4873.
References
1. Jager, H. I., D. L. DeAngelis, M. J. Sale, W. VanWinkle, D. D. Schmoyer, M. J. Sabo, D. J. Orth & J. A. Lukas. "An individual-based model for smallmouth bass reproduction and young-of-year dynamics in streams." Rivers. Vol. 4, No. 2, pp.91-113 (1993).
2. Van Winkle, W., H. I. Jager, S. F. Railsback, B. D. Holcomb, T. K. Studley & J. E. Baldrige. "Individual-based model of sympatric populations of brown and rainbow trout for instream flow assessment: model description and calibration." Ecological Modelling. Vol. 110, No. pp.175-207 (1998).
3. Chambers, R. C. "Phenotypic variability in fish populations and its representation in individual-based models." Transactions of the American Fisheries Society. Vol. 122, No. pp.404-414 (1993).
4. Jager, H. I. "Individual variation in life history characteristics can influence population extinction risk." (in prep.).
5. Domeier, M. L. a. P. L. C. "Tropical reef fish spawning aggregations: defined and reviewed." Bulletin of Marine Science. Vol. 60, No. 3, pp.698-726 (1997).
6. Cochnauer, T. G., J. R. Lukens & F. E. Partridge (eds.) Status of white sturgeon, Acipenser transmontanus, in Idaho Dr W. Junk, publishers, Dordrecht, The Netherlands, 1985.
7. Jager, H. I., W. V. Winkle, K. Lepla & J. Chandler. "Are white sturgeon dammed? -- Population viability of white sturgeon in a fragmented river habitat." (submitted to Conservation Biology).
8. Schlosser, I. J. "Stream Fish Ecology: A Landscape Perspective." Bioscience. Vol. 41, No. pp.704-712 (1991).
9. Semakula, S. N. & P. A. Larkin. "Age, growth, food, and yield of the white sturgeon (Acipenser transmontanus) of the Fraser River, British Columbia." Fish. Res. Bd. Canada. Vol. 25, No. 12, pp.2589-2602 (1968).
10. Lomnicki, A. Population Ecology of Individuals. Princeton, NJ: Princeton University Press,1988.
11. van Noordwijk, A. J., Conservation Genetics. Basel, Switzerland: Birkhauser Verlag, Loeschcke, V., Tomiuk, J. & Jain, S. K. (eds.) (1994) pp. 131-147.
12. Rice, J. A., J. T. Miller, K. A. Rose, L. B. Crowder, E. A. Marschall, A. S. Trebitz & D. L. DeAngelis. "Growth rate variation and larval survival: inferences from an individual-based size-dependent predation model." Canadian Journal of Fisheries and Aquatic Sciences. Vol. 50, No. 1, pp.133-142 (1993).
13. Lande, R. & S. Shannon. "The role of genetic variation in adaptation and population persistence in a changing environment." Evolution. Vol. 50, No. 1, pp.434-437 (1996).
14. Gilpin, M. E. & M. E. Soule, Conservation biology: the science of scarcity and diversity. Sutherland, MA: Sinauer Associates, Soule, M. E. (ed.) (1986)
15. Hindar, K. (ed.) Alternative life histories and genetic conservation Birkhauser Verlag, Basel, Switzerland, 1994.
16. Kareiva, P. M., J.G. Kingsolver, and R.G. Huey (ed.) Biotic Interactions and Global Change Sinauer Associates Inc., Sunderland, MA, 1993.
17. Jager, H. I., W. V. Winkle & B. D. Holcomb. "Would hydrologic climate changes in Sierra Nevada streams influence trout persistence?" Transactions of the American Fisheries Society. Vol. 128. pp. 222-240. (1999).