
Last modified Saturday 21st June 1997
EUROPE east to 40° E (the Urals), and including Asia Minor
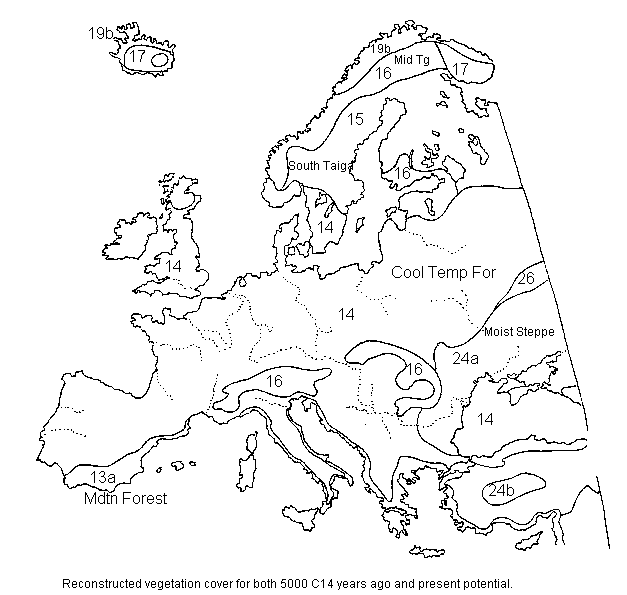 Europe 5,000 14C years ago and present-potential
Europe 5,000 14C years ago and present-potential
Also refer to Eurasia (for Europe east of Black Sea).
VIEW NEW SET OF PALAEOVEGETATION MAPS (PRELIMINARY) AT MORE FREQUENT TIME INTERVALS
18,000 years ago (last glacial maximum or LGM) map.
List of References (separate document)
Key to the vegetation classification system used in the atlas
Distribution of sites used towards reconstructing the LGM palaeovegetation distribution for Europe
Sea level. For cartographic convenience, shorelines are shown at the -150m bathymetric contour. Sea level was probably in fact 120 - 140 m lower than at present (H. Faure pers. comm. June 1992), and would roughly correspond to the -150m contour. The Black Sea was also somewhat lower; the Caspian Sea was higher for reasons which do not seen clearly explained in the literature. The shorelines of the lakes shown here are from Velichko & Kurenkova (1990) and E. Zelikson (pers. comm., May 1994).
Ice extent. The extent of the main European ice sheet is taken from Denton & Hughes (1981), modified according to more recent data used to produce a map published by Velichko & Isavea (1992). Note that there remain uncertainties on the precise position of the ice margins at 18,000 years ago (e.g. see discussion on p.49 in Dawson, 1992). Possibly, an ice-free area would have been present in what is now the North Sea, completely separating the British ice sheet from the Scandinavian one (Sejrup et al. 1987). Ice may have extended further east into Siberia than the limit shown on this map, but the ice extent shown here agrees broadly with the opinion now apparently held by most experts. Some maps showing ice extent that are presented by various authors in Soffer & Gamble (1990) also appear to show the ice extending further south in central Europe; it is not clear whether these are due to errors in cartography or differences in opinion about the maximum extent of the last glaciation.
Large ice caps were present over the Alps, and over parts of the Pyrennes, and smaller ice caps were also present on various other mountainous areas around Europe (e.g. as shown in the map by Grichuk 1980, and by Denton & Hughes 1981). As with other parts of the world, the concept of a true 'glacial maximum' depends on the individual location; in the Pyrennes, ice extent was apparently less at 18,000 14C years ago than it had been several thousand years earlier (Denton & Hughes 1981), possibly due to reduced precipitation at the LGM.
The LGM vegetation in general. The long tradition of study of the Quaternary in Europe has ensured a relative abundance of data for this region. The earliest mapping attempts which suggested that the full-glacial environment of Europe had been moist and extensively forested have now been superseded, and it is apparent that the LGM environment of Europe was much more barren of vegetation than previously thought. Only the maps of Grichuk (1992) persist in showing large areas of forest in southern Europe. However, even here the intention is to indicate biogeographical 'refuge' areas rather than actual vegetation cover (Zelikson pers. comm. 1994), a fact which has frequently been overlooked by those who consult his maps. Other maps based on compilation of a range of data sources (e.g. Frenzel 1992, Velichko & Isavea (1992) concentrate on the actual LGM vegetation cover, and agree in suggesting a virtually treeless semi-arid environment in both northern and southern Europe. This view would certainly seem to be borne out by the mass of evidence from across the region, reviewed below.
The Mediterranean region at the LGM. From pollen evidence, there appears to have been very little closed woody vegetation in the Mediterranean countries at around the LGM. From mainly biological indicators, summer temperatures all across southern Europe seem to have been about 6 to 8°C cooler than at present, with winter temperatures 10 to 12°C lower (Frenzel et al. 1992).
A semi-desert steppe predominant in southern Europe. Throughout southern Europe, the predominant picture for the LGM is of an arid, semi-desert like steppe perhaps similar to the cool and arid montane steppes found today in the Pamirs of southern Russia (Tallis 1990 p.204) but also resembling the driest and most eroded present-day landscapes of south-central Spain (G. Jalut, pers. comm. June 1993). The generally sparse nature of the vegetation cover is indicated by many pollen cores from Spain across to Turkey which date from the LGM and/or the Late Glacial (e.g. Wijmstra 1969, Frenzel 1973). The drought-tolerant weedy plant genus Artemisia was almost always an important component of the pollen spectrum. Species of this genus are now found in abundance in such dry, open habitats as builders' rubble and on the central Asian deserts, but some of them also occur in fairly mesic conditions as colonisers of open ground in fields and on river banks. As the fossil pollen record does not allow one to distinguish between species of Artemisia , it is not possible to know exactly which types were present. All that can be said with confidence is that there was a high proportion of open ground (presumably due to aridity) and thus a sparse vegetation cover (Tallis 1990 p.199-204, E. Zelikson, May '94). Chenopods and grasses were also important elements of the vegetation; the chenopodeaceae seeming to indicate the salty soils characteristic of semi-desert conditions (Frenzel 1973). However, there is often a small component (less than 10%) of Pinus in the pollen diagrams, suggesting the persistence of thinly scattered trees or small pockets of open woodland where local soil moisture levels allowed this.
Some areas of woodland present. In southern Europe, some areas of open pine (Pinus) and birch (Betula) woodland seem to have existed in the plains of northern Italy and to the south-east of the Alps, although these were not extensive (C. Tzedakis, August 1995). Scattered pockets of woodland or isolated trees occurred through the mid-altitudes (e.g. with a belt about 500m above [present] sea level in Northern Greece) of the southern Balkans and Apennines, according to reviews of the pollen evidence by van Zeist & Bottema (1982), Bennett et al. (1990) and Willis (1994). S. Bottema (pers. comm. April 1996) suggests from the pollen diagrams that the refugia for deciduous and needle-leaved species were mainly on the western side of the Mountains of Greece.
In the Balkans, the several scattered pollen sites indicate that the predominant vegetation was an open Artemisia steppe with a small component of Pinus, but with many other tree taxa present and locally abundant at mid elevations (around 500m altitude), possibly due to the orographic rainfall and reduced evaporation supplying just enough moisture to enable them to survive.
In Iberia, scattered patches of pine woodland may have been present in valleys and other sheltered sites in the mountains of Spain (Turner & Hannon 1988), showing up to some extent in the pollen diagrams. Yet although a Pinus and Quercus element was present, it seems to have been only locally important. For the most part, the LGM vegetation of Spain and Portugal seems to have been drought-tolerant, open and semi-desert or steppe-like, with abundant Artemisia, and also chenopods and grasses. Several cores scattered around the Iberian peninsula and off the south-west coast all indicate the same general vegetation type (Hooghiemstra et al. 1992). The most recently published core confirms the same picture of an almost treeless cold-tolerant, arid steppe for the region close to the Mediterranean, inland from Barcelona in the north-east of Spain (Pérez-Obiol & Julià 1994). In the south-west of Spain, the vegetation on the exposed continental shelf areas seems to have been sparse enough to allow extensive dune mobility at the LGM (Zazo pers. comm., May 1994), even well inland from coastal areas. In south-eastern Spain, sparse vegetation cover and semi-arid conditions punctuated by sudden storms are indicated by slope wash deposits which are thought to date from the last glacial (Harvey 1984).
Paradoxically high lake levels. Paradoxically, given the overwhelming pollen and sedimentological evidence of aridity, lake levels in several parts of southern Europe were about as high as their Holocene values (Harrison et al. 1993, 1996). It has been suggested on the basis of GCM modelling that high winter rainfall, in the form of intense storms with high runoff, was able to fill the lakes without making much moisture available to the plants (Harrison et al. 1993). Lower year-round temperatures would have suppressed evaporation from these lakes.
Central and eastern Europe.
Dry cold steppe with scattered pockets of trees.
Mean annual temperatures from scattered geomorphological evidence are estimated to have been around 13-17°C lower than at present (Frenzel 1992 a,b, Dawson 1992). The greatest cooling was in winter (February temperatures 12 to 20 degrees lower), but biological indicators suggest that August mean temperatures were around 8 to 10°C cooler than today.
These temperatures translate into an estimate that at about the latitude of south-central Germany and north-western Ukraine, August mean temperatures were (at most) 10-11 Celsius, comparable with much of the northern coast of Siberia at present. Winter (February) mean temperatures would have been at least as low as -19 Celsius in central Germany and -27 Celsius across most of the northern Ukraine (comparable with much of northern and central Siberia at present).
Most pollen cores for eastern and central Europe show a sparse element of cold-tolerant trees at around the time of the LGM (Soffer 1990). These were trees probably existing either as isolated pockets or as scattered individuals in an open landscape (Kowzlowski 1990, Zelikson pers. comm.). Further east, in the Russian Plain, gallery forest or woodland seems to have existed along river valleys, in a vegetation that was predominantly open steppe or steppe-tundra (e.g. landscape map on p.234 in Soffer 1990, & Velichko pers. comm.).
Soffer (1990) also shows a belt of forest-steppe (open vegetation with isolated trees or pockets of trees with a herbaceous ground layer) to the north and east of this area, as do Velichko & Kurenkova (1990), whose map shows a steppe and open forest-steppe landscape south of the steppe-tundra in Eastern Europe. The term forest-steppe is perhaps misleading, as it tends to invoke the picture of a major woody component in the landscape, which was certainly not intended by the authors (Velichko pers. comm. May 1992). Groves of cold-resistant trees (Pinus, Betula, with some Larix and Picea) were present, but as isolated pockets in sheltered spots, in a mainly treeless landscape. Knowledge of the vegetation at around the LGM period is based on pollen grains isolated from loess sections, although radiocarbon dating is not usually possible. Generally, the indications are of an open-ground mainly herbaceous flora rich in Artemisia.
In the vegetation scheme being used here for the production of the QEN maps, a much higher woody coverage (>20%) would be necessary for this to be categorised as forest steppe. In the maps of Grichuk (1992), Frenzel (1992) and Velichko & Isayeva (1992), essentially the whole of eastern Europe is categorised as 'periglacial steppe' or 'periglacial tundra' with a sparse, herbaceous vegetation cover.
Loess-ridges, of windblown dust forming dune-like structures, are widespread across SE Poland, Belorussia, Ukraine, Romania and Bulgaria (literature discussed and summarised by Wells 1992 p.687) and they seem to reinforce the view that there was only a sparse vegetation cover in these areas in order for the surface deposits to have been so mobile. Furthermore, there seems to have been little or no soil development within the many loess sections that span the LGM period in Russia and the Ukraine, indicating that conditions were dry with limited plant growth and biological activity (E. Zelikson, pers. comm. May '94). Interpretation of biogenic and non-biogenic calcite structures from the central and east European loess soils of the last glacial is further seen as evidence for an arid environment (Becze-Deak et al. 1995). However, it is thought to have been the presence of at least some (albeit sparse) vegetation which allowed this loess to become stabilised.
Zoological evidence from around the time of the LGM also indicates a predominantly cold, semi-desert or dry steppe environment in which the loess was often deposited (Soffer 1990, Baryshnikov & Markova 1992, E. Zelikson pers. comm., C. Kreminetski pers. comm.). Precipitation, presently around 600mm, may have been only about 60-120mm per year at this time even without allowing for the temperature lowering (Soffer 1990). However, humans do seem to have been present at the LGM, as several well-dated encampment sites have been found for the Russian Plain at this time.
Some areas of woodland near to the Carpathians. According to Soffer (1990), pine (Pinus) (with birch and spruce) woodlands may have existed to the south and east of the Carpathian mountains.
There is pollen and pedological evidence (published in Russian) for at least some areas of relatively mesic steppe, probably with a dense vegetation cover and full soil development, in certain areas to the south of the Carpathians, especially over Rumania, shown on the maps of Grichuk (1980). Yet, in the plains of Hungary to the south-west, and to the north of the Carpathians drifting sand sheets and parabolic dunes seem to have been extensive at around the time of the LGM (Borsy 1987, Wells 1992 p.687) indicating that there would have been only a sparse vegetation cover in these other adjacent areas. This seems to restrict the possible area of woodland to an area immediately to the south-west of the Carpathians.
North-western Europe.
Treeless, dry arctic conditions. To the north and west, in Germany, northern France, the Low Countries and Britain, the landscape appears to have been completely lacking in trees, as arboreal taxa do not appear in the pollen records for these areas (Weniger 1990). For example, at Grande Pile (47 N) in central France, tree pollen is present but only at very low abundance (with the open ground herbaceous genus Artemisia being dominant) (Guiot et al. 1989). The Les Echets core (Guiot et al. 1989) shows more (20%) tree pollen but this may be reworked from older layers. From Grande Pile, a 9.5°C LGM depression in mean annual temperature is suggested (Guiot et al. 1989). Estimates based on geomorphological indicators suggest a mean annual temperature depression of at least 15-17°C all across the area ranging from central England down to south-western France (e.g. climate reconstruction maps in Frenzel, Pecsi & Velichko 1992, and data summarised by Dawson 1992). Summer temperatures across most of NW Europe were around 10-12 °C cooler than today. Permafrost extended down to southern France, just north of Bordeaux, and into the uplands of northern Province (Dawson 1992).
The areas just to the south of the main ice sheets in north-west Europe seem to have had only very sparse vegetation, based on animal and plant fossil evidence and pedological evidence of very cold and dry conditions (e.g. as summarised in West 1978). Where deposition continued through the LGM, there seem to have been very low pollen influx values, indicating little or no vegetation across this region. Dune activity during the last glacial seems to have been quite widespread in England, northern France and the Low Countries, and eastwards across Germany, Czechoslovakia and Poland (reviewed by Wells 1992 p.687). This reinforces the view of arid and sparsely vegetated conditions.
Generally, the picture is of something approaching polar desert close to the ice sheets, but with slightly more vegetation cover (but still very open) from northern France southward.
North-eastern Europe.
Moistest close to ice sheet, dry further south. In contrast, pollen evidence from close to the southern edge of the ice sheet in Russia indicates relatively moist conditions (although still much drier than most present-day tundra), in a belt extending for several hundred kilometres away from the ice sheet margins (Grichuk 1980, Zelikson pers. comm. 1994). In general, the LGM vegetation in this belt seems to have resembled tundra in many aspects of its floristic composition, although it would appear to have been rather drier than most present-day tundra, with the overall ground cover by vegetation of perhaps around 50%, as a rough estimate (E. Zelikson, May '94). A peat layer would seem to have been absent, with only relatively poor soil development and low organic matter contents being recorded from buried soils in this area.
To the south of this tundra-like belt the pollen diagrams indicate that there was a drier, rather more sparsely vegetated 'peri-glacial steppe' or 'steppe-tundra', with many elements of what is presently dry steppic vegetation. As mentioned above, there was a high relative abundance of such open-ground and semi-desert species as Artemisia, indicating that the percentage ground cover by vegetation would have been low. E. Zelikson (pers. comm.) suggests that the ground cover by plants was at most 50%.
There are almost no remains of peat deposits of any sort within either the 'tundra-like' or the 'steppe-like' belts of the steppe tundra of the late glacial. The only organic horizons that date to around the LGM are occasional thin layers of twigs and other coarse plant debris (E. Zelikson pers. comm., May 1994) .
Caucuses.
Scattered pockets of woodland. The Caucuses is thought to have been a glacial refuge for many temperate tree taxa, although there is little direct evidence of these trees surviving there during the LGM. In the Colchis region of Southern Russia, along the eastern shores of the Black Sea and the high ranges of the Caucuses, pollen evidence indicates that there were scattered pine and birch forests but broad-leaved trees were probably very localised in distribution (Tumajanov 1971). In contrast to the large area of surviving forest suggested in the maps Grichuk (1980, 1992), Velichko & Kurenkova (1990) and Frenzel (1992) also show only small areas of closed temperate forest surviving in the lowlands of the southern Caucasus, with dry steppe and open woodlands or localised woody pockets prevailing. The map by van Zeist & Bottema (1988) similarly suggests that there was virtually no temperate forest present in the main Caucasus region at the LGM. It is not clear from these sources what evidence is used to justify this choice of vegetation types or to what extent more recent research on this area has been published.
The shorelines of the Black Sea and Caspian Sea were different from today due to changes in the water supply and evaporation rates, and possibly also tectonic changes (Velichko & Kurenkova 1990). The Black Sea was shallower than today, whereas the Caspian Sea was somewhat deeper and more extensive (Velichko & Kurenkova 1990).
Asia minor.
Much less woody cover. At sites scattered through Turkey, Syria and eastward into Iran, there are a number of pollen-bearing cores that date back to about the LGM. An LGM-to-16,000 14C y.a. map presented by van Zeist & Bottema (1988) indicates a considerable reduction in woody vegetation and an expanded area of steppe and semi-desert steppe in the upland areas of Turkey, northern Syria and western Iran, relative to the present-natural state. Open woodland or wooded-steppe is suggested as having survived over much of western, southern and eastern Turkey. Mesic deciduous forest is reconstructed for the north, as a thin band along the shore of the Black Sea, and there is also woodland along the southern coast of the Caspian Sea, although no pollen localities are cited as supporting the existence of woody cover in these areas. Woodland is thought likely to have been present along the western Levant, but of a more open character than the late Holocene present-potential vegetation.
Lake-level evidence from three sites in Turkey (Lake Van, Konya Basin and Lake Urmia) suggests that at least in terms of rainfall, LGM conditions might not have been much drier than present (Landmann pers. comm. April 1997). The aridity evident in vegetation records may have come from a pattern of sudden winter storms, filling the lakes, but not making water available to the vegetation. From the lake record, maximum aridity seems to have been reached about 16,000-15,000 14C y.a.
A note on the nature of the steppe-tundra vegetation. Most of the open vegetation types that predominated in northern Europe at the LGM are described by palaeoecologists as 'steppe-tundra'. The steppe-tundra was a largely herbaceous vegetation type that seems to have extended in broadly the same form across much of Eurasia, Beringia and parts of North America. For example, according to Guthrie's (1990) maps, this vegetation category includes many of the areas referred to above as 'steppe' or 'periglacial steppe' by eastern European authors. As a factor in common, floristic composition seems to have no close present-day analogue, as it combined species that presently live in quite distinct environments (Hopkins et al. 1982). Artemisia was always a major component, together with grasses and chenopods. Other plants that were less abundant (but nevertheless occurred) included species of present-day moss lichen tundra (Cyperaceae), dry fellfield (Armeria, Cerastium, Rumex), arctic-alpine communities (Saxifraga), cool steppe (Kochia), saline communities (Plantago maritima, Sueda) and dwarf-shrub communities (Juniperus, Salix) (Schweger 1982, Tallis 1990). It is possible that in the LGM steppe-tundra there was a small-scale mosaic of a wide range of plant communities in any one area (Schweger 1982). The closest modern analogues appear to be scattered patches of what is known as 'tundra-steppe' on steep south-facing slopes in north central and north eastern Siberia (Yurtsev 1982) and in the Brooks Range of Alaska (Schweger 1982), but this modern vegetation seems to have a proportion of moisture-requiring species (e.g. Eriophorum)which were absent or much less common in the LGM vegetation (Tallis 1990) although this might partly reflect the fragility of these pollen types in terms of preservation (E. Zelikson pers. comm. May 1994).
Guthrie (1982) has claimed that the very varied mammal fauna found on the full glacial steppe-tundra belt indicates that it must have been a biologically very productive vegetation type. However, most palaeoecologists seem to reject this view in favour of a mosaic of tundra, steppe and scrub communities of lower productivity and with a sparse overall ground cover (Schweger 1982, Tallis 1990). Tallis notes, for example, that radiocarbon dates of mammoth finds across the whole steppe-tundra belt reach a low point at around the LGM, suggesting perhaps that the vegetation did not support an especially dense population of these herbivores (though if conditions were arid, this might itself be expected to decrease the possibility of preservation by burial in water-borne sediments). It is suggested, partly by Guthrie (1982) amongst others, that it was the mosaic nature of the steppe-tundra vegetation rather than high biomass / productivity that allowed a range of herbivores to coexist at the time.
Europe 8,000 14C years ago (early Holocene)
Vegetation communities of the region in general. Huntley (1990) has published maps of broad changes in the vegetation types across Europe during the late glacial and Holocene, based on a wide spread of pollen data from cores, and numerical analysis of this. The map here is partly based on Huntley's 8,000 14C y.a. map, which shows a much greater extent of Betula and Pinus-dominated woodland in central and eastern Europe, relative to the 5,000 years ago and present-natural situation. We have attempted to fit the categories that he has mapped into the standard vegetation scheme used here. However, consultation with specialists and particular literature sources for sub-regions of Europe have shown what appear to be significant inaccuracies in Huntley's maps (see below). The 9,000 and 6,000 years ago maps presented by Huntley & Prentice (1993) (also on the basis of numerical analysis of pollen data) seem more realistic as a general summary of the lower Holocene, but still possibly subject to certain inaccuracies.
Lack of agricultural impact. Agriculture seems to have been present in Turkey and eastern Greece by around 8,000-9,000 14C y.a., but its impact on vegetation seems to have been very localised (Bottema et al. 1993, Willis & Bennett 1994). Elsewhere in Europe, the only human communities present seem to have been hunter gatherers.
Northern Europe.
The last remnants of the Scandinavian ice cap may have disappeared at or around 8,000 14C years ago (Denton & Hughes 1981, Dawson 1992), but in the absence of any published maps no attempt is made to show these here. Both summer and winter temperatures were higher than present, though perhaps not as warm as during the period 7,000-5,500 14C y.a.
Mainly forested in northern & central Europe, but slightly drier. In Northern Europe and Scandinavia, climatic conditions seem to have been broadly similar but slightly drier than at present, on the basis of the lake level evidence (e.g. various sources cited and summarised in Harrison et al. 1993). A few areas of steppe and steppe-woodland seem to have persisted until around 8,000 years ago in scattered areas in central Europe (Huntley 1990) on sandy soils, perhaps due to a drier more continental climate at that time (Starkel 1991). However, the overall picture is that central Europe was heavily forested by 8,000 years ago.
Peat build-up just beginning. Peat build-up in Finland and the British Isles had already started by around 9,000 14C years ago (evidence summarised by Tallis 1990). However, the extent to which the later peat cover had already developed at 8,000 years ago is not expressed in quantitative terms by his diagrams. In NW parts of European Russia, peat build-up also seems to have begun around 8,000-9,000 years ago in most sections studied (data published in Russian: pers. comm. C. Kreminetski, Acad. Sci., Moscow. May 1994).
Relatively open forests in NE Europe. In European Russia, the abundance of Betula (at around 50% of total pollen) (Huntley 1988, 1990, 1992, & E. Zelikson, Acad. Sci. Moscow, May 1994 pers. comm.) across a wide swathe of northern and eastern Europe in the early Holocene suggests that many of the more northerly forests were still relatively low in biomass, possibly due to ongoing successional processes related to soil or migrational lag.
Steppic belt not forested. Huntley (1992) and Huntley & Prentice (1993) suggest that what is presently the steppic region of southern Russia, Ukraine and Bulgaria seems to have been largely forested during the lower and mid Holocene, with Picea, Betula and Pinus . However, E. Zelikson (pers. comm., May 1994) is strongly of the opinion that this area was not forested, and that although Betula and Pinus were present, they were a very minor element in the landscape (and Picea was not present at all). Instead, Zelikson suggests the pollen diagrams, buried soils and zoological evidence from this region show dominance by dry steppic herbaceous species, typical of present-day open steppes.
More forest in eastern Caucuses. Pollen records recently obtained from the eastern Caucuses mountains of Georgia show that during the lower Holocene the mesic deciduous forest was considerably more widespread than now, in areas that are presently only xerophytic scrub-woodland (Gogichaishvili 1995). A decline in forest extent began by the mid-Holocene, perhaps due to human activity (Gogichaishvili 1995).
The Mediterranean.
Moister than present in Mediterranean region? In the Mediterranean region, lake levels across from southern France to Greece seem to have been higher than present around 8,000 years ago, indicating moister-than-present conditions (Harrison et al. 1993).
Deciduous forests covering Spain and elsewhere in Mediterranean region. In the southern Iberian Peninsula, various land and sea pollen cores indicate a change from steppe-like vegetation to deciduous oak forest at around 10,000-9,000 14C years ago (Hooghiemstra et al. 1992), with the maximum of oak pollen being around 9,100-8,400 years ago. This evidence seems to contradict Huntley's (1990, 1992) map interpretation of steppe and steppe-woodland for 8,000 years ago. Also, in the extreme north-east of Spain, a lake pollen core indicates that deciduous oak (Quercus) forest was dominant - together with a small amount of evergreen oak (Q. ilex) and pine (Pinus) - at 8,000 years ago (Pérez-Obiol R. & Julià R. 1994.).
Likewise, in the present Mediterranean scrub region of southern France, pollen cores indicate closed deciduous temperate forest at 8,000 yr. ago. (Laval et al., 1991). Huntley (1990) depicts most of the northern Mediterranean fringe (e.g. Italy) as deciduous broad-leaved forest at 8,000 years ago, but with a considerable area of Quercus-Pinus sclerophyll woodland occurring in Northern Greece and along the Balkans. However, S. Bottema (pers. comm., April 1996) is of the opinion - based on his interpretation of the pollen evidence - that the eastern Mediterranean (Greece, Turkey and the Levant) had essentially no Mediterranean Forest/Mediterranean Scrub during the early-to-mid Holocene, and that 'Mediterrean vegetation' would still not exist today without human intervention. Instead he suggests that the vegetation even near to the coasts would have been dominantly deciduous oak forest, but with a lower story of Kermes oak, Phillyrea, Arbutus etc. on dry rocky pockets. If so, the zones of Mediterranean Forest in the eastern Mediterranean marked on the Holocene maps shown here are inappropriate and should be replaced by temperate mixed/deciduous forest.
Although deciduous forest was apparently predominant in Spain during the early Holocene, pollen suggesting evergreen oak and pine forest turns up in a lake in the Ebro Basin in NE Spain (currently one of the most arid parts of Spain) between 8,600 and 7,200 years ago (though the lake level temporarily fell between 8,200 and 7,600 years ago, interrupting the pollen sequence, perhaps indicating drier conditions than this at 8,000 years ago) (Davis 1995).
Asia minor. Expanded woody vegetation, but less extensive than late Holocene natural. For Turkey and the surrounding areas, the map given by van Zeist & Bottema (1988) indicates an expansion in woody vegetation relative to the LGM, although it does not seem to have been as extensive as later in the Holocene. A belt of woodland and (towards the east) forest-steppe seems to have extended eastwards across the mountains of northern Iraq and western Iran. This interpretation is based largely on pollen evidence, from sites scattered through this region. Roberts & Wright (1993) also present Holocene maps of this region, which are in agreement with this general interpretation.
Europe 5,000 14C years ago and present-potential
Warmer climates in northern Europe/Scandinavea. There are various sources of evidence for warmer summer and winter temperatures across northern Europe during the mid-Holocene. These indicators do not all agree on the same time slices or intervals for maximum warming, but in general it seems that the period between 7,000 and 4,000 14C years ago includes the warmest part of the Holocene in most areas. The diverse data have been been summarized into climate map reconstructions, indicating about a 1 deg.C warming in mean July temperature at 50 degrees North, a 2 degrees warming above 60 degrees North, and 3-4 degree warming in above about 65 degrees North (Klimanov et al. 1992). A winter warming of about 2 degrees is suggested for most of north-west Europe (Klimanov & Velichko 1992). The available indicators do not all agree on the same time slices or intervals for maximum warming, but in general it seems that the period between 7,000 and 4,000 14C years ago includes the warmest part of the Holocene in most areas.
Agriculture still localised at 5,000 years ago. By 5,000 14C years ago, agriculture had spread to most parts of Europe. Generally, however, there seems to have been no substantial effect of agriculture on pollen diagrams in Europe (nor even in the agricultural 'cradle' of the Near East) before 4,000 14C years ago (e.g. Tallis 1990, M.K. Jones pers. comm., Jan 1992). There is increasing evidence that the Europe-wide 'elm decline' of around 5,000 years ago was the effect of a short-lived climatic deterioration and not a sudden increase in agriculture and herding as had previously been thought (Rackham 1980, Peglar 1993). Even in Greece, significant soil erosion due to deforestation does not seem to have occurred until after 4,000 years ago (M.K. Jones, pers. comm. Feb. 1992, S. Bottema pers. comm. April 1996). Crete was an exception, with pollen evidence suggesting loss of the original deciduous forest cover and its replacement by maquis by about 5,000 years ago (Unpublished data: O. Rackham, pers. comm. 1992), and likewise some parts of the English chalklands may have been largely cleared for agriculture by 5,000 years ago (Evans 1975). However, these localised (and often transitory) occurrences should not be allowed to distract from the overall pattern. Willis & Bennett (1994) argue on the basis of their review of pollen evidence from around Europe, that agricultural impact on the vegetation almost everywhere was negligible until after 4,000 years ago.
Steppic zone existing. In eastern Europe, Huntley (1990) appears to suggest in his map that the present-day steppic region of Ukraine and Southern Russia was mainly forested, partly with pine-birch and partly with mixed conifer-hardwoods. However, E. Zelikson (pers. comm., May 1994) is sure from her own work on this region that it was steppe vegetation much as today's, with only very scattered woody elements in the most favourable microsites. She suggests that these localised woody pockets may have biased the pollen records that Huntley is interpreting.
Expansion of mediterranean forest. By 5,000 yr. ago., evergreen mediterranean vegetation was more widespread through Greece and Italy than at 8,000 yr. ago., replacing the deciduous forest in localized areas on limestone soils. In southern France, however, pollen evidence indicates that closed deciduous forest was present in areas that are now evergreen scrub (Laval et al., 1993), although with some indications of sporadic clearance. Widespread clearance and replacement by maquis and garrigue in southern France seems to have occurred only after around 2,000 yr. ago. (Laval & Medus 1989, Laval et al. 1991).
In the Ebro Basin of NE Spain, by 5,000 yr. ago the pine/evergreen oak forest cover was decreasing (possibly due to increasing climatic aridity; there are no charcoal or cultivation indicators at this time period), and lake levels were lower than during the earlier Holocene (Davis 1995). B.A.S. Davis (pers. comm. Nov 1995) notes that arid Pinus halepensis forest was more widespread at lowland sites than earlier or later in the Holocene, and lake levels were low with a hiatus (indicating drying-up of lakes) at some stage during the mid-Holocene (Reed 1995 and Fatima 1995).
Similar to present-natural in Turkey. The map of van Zeist & Bottema (1988) for 4,000 years ago (based on several scattered pollen-bearing cores in Turkey and Iran), indicates that the vegetation in Turkey included a wide band of temperate forest in the north and south, with a belt of woodland extending eastwards across the mountains of northern Iraq and western Iran. It seems that in general the vegetation was similar to its (hypothetical) late Holocene present-natural state, and without extensive anthropogenic deforestation by 5,000 years ago.
List of References (separate document)
Names and current addresses of QEN participating experts (named in the text above) who have made direct contributions to this work on Europe:
B.A.S. Davis, University of Newcastle, UK. e-mail; B.A.S.Davis@newcastle.ac.ukS. Bottema, Vakgroep Archaeologie, Poststraat 6, 9712 ER, Groningen, Netherlands. Bottema@let.rug.nl
H. Hooghiemstra, Hugo de Vries Laboratory, University of Amsterdam, Kruislaam 318, 1098 SM, Netherlands.
C. Kreminetski, Institute of Palaeogeography, Russian Academy of Sciences, Staromonetny per 29, Moscow 109017, Russia.
M.K. Jones, Department of Archaeology and Anthropology, University of Cambridge, Cambridge CB2 3EA, UK.
G. Landmann, Intitut fuer Biogeochimie und Meerschemie, Universitaet Hamburg, Jungiusstr. 6, 20355, Hamburg, Germany.
H. Laval, URA CNRS 1152, Palynologie, Faculte des Sciences et Techniques de Saint Jerome, C.541, F-13397, Marseille Cedex 13, France.A. Parker, School of Geography, University of Oxford, 1 Mansfield Road, Oxford OX1 3TB, UK.
O. Rackham, Corpus Christi College, University of Cambridge, Cambridge, UK.
N. Roberts, Department of Geography, Loughborough University, UK.
F.A. Street-Perrott, Department of Geography, University of Swansea, Swansea, UK.
C. Turner, Sub-Department of Quaternary Research, Department of Plant Sciences, University of Cambridge, Cambridge CB2 3EA, UK.
C. Tzedakis, Sub-Department of Quaternary Research, Department of Plant Sciences, University of Cambridge, Cambridge CB2 3EA, UK.
A.A. Velichko, Institute of Palaeogeography, Russian Academy of Sciences, Staromonetny per 29, Moscow 109017, Russia.
E. Zelikson, Institute of Palaeogeography, Russian Academy of Sciences, Staromonetny per 29, Moscow 109017, Russia.
 Europe 18,000 14C years ago
Europe 18,000 14C years ago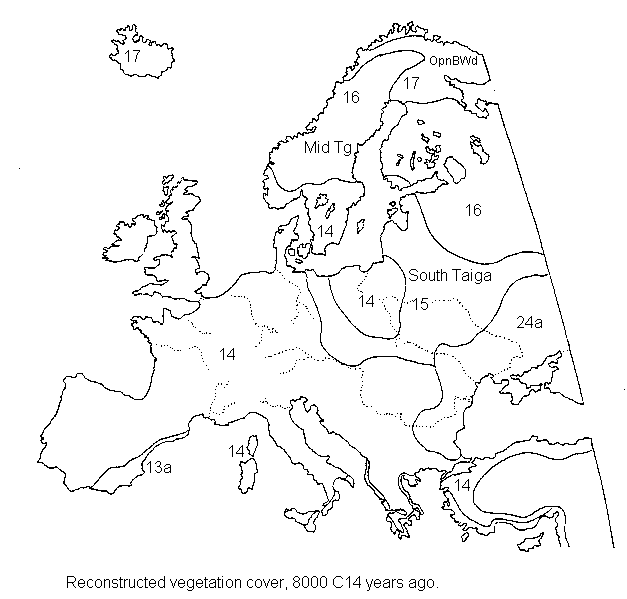 Europe 8,000 14C years ago
Europe 8,000 14C years ago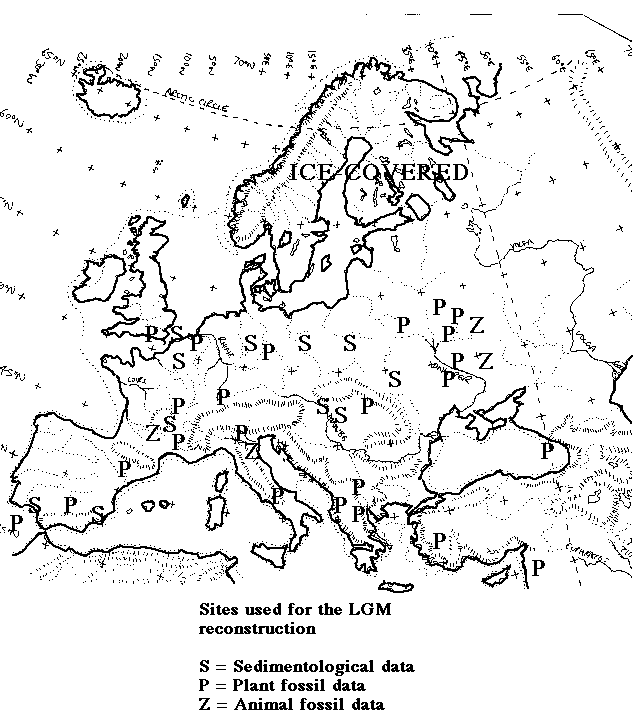{kind=link}