Last modified 16th April 1998
AFRICA
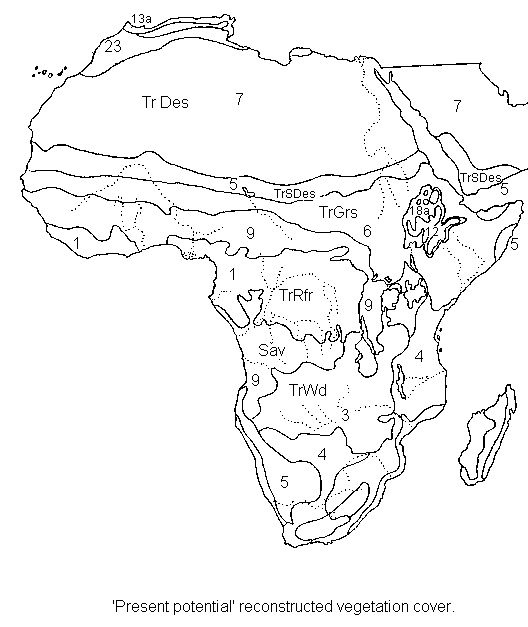 Africa present-potential vegetation
Africa present-potential vegetation
VIEW NEW SET OF PALAEOVEGETATION MAPS (PRELIMINARY) AT MORE FREQUENT TIME INTERVALS
Distribution of sites used towards reconstructing the LGM palaeovegetation distribution for Africa
The 'glacial maximum'. There is some ambiguity in the concept of the 'glacial maximum' as applied to the tropics. It may be that in parts of Africa, the coolest temperatures and maximum ice extent in mountain glaciers occurred earlier or later than the maximum ice extent at higher latitudes. For example, the mountain glaciers of the Ruwenzori Mt.s (Uganda) may have reached their maximum extent around 15,000 years ago (Livingstone 1980, 1993).
The Levant Goodfriend & Margaritz (1988) conclude that the Levant at the LGM generally had rather more arid conditions than at present. Two pollen localities, one in northern Israel and the other in northern Lebanon, suggest a band of woodland or forest-steppe running along the uplands parallel to the coast, and reaching perhaps 50 km inland. Note that this area, though now barren as a result of deforestation and overgrazing, seems to have supported a moister-climate closed forest (rather than the LGM open woodland/forest-steppe) vegetation up until the late Holocene (Goodfriend & Margaritz 1988). On the basis of these same cores, Bar-Yosef likewise concludes that the coastal ranges of Syria, Lebanon and Israel apparently received enough winter precipitation to be covered in forests, which became more open toward the south. He suggests that only a narrow strip of parkland vegetation stretched along the Transjordanian Plateau.
Conditions also seem to have been colder and drier over the rest of the region, with zoological and sedimentary evidence suggesting that the steppic belt was reduced and the Saharo-Arabian desert belt extending further westwards into Syria and Jordan (Bar-Yosef 1990).
Drier in the Sinai-Negev area. Bar-Yosef (1990) notes that the arid Saharo-Arabian belt was expanded northwards in the Sinai and Negev region at the LGM. There was erosion and burial of loess by sand sheets at the LGM itself, indicating considerably more arid conditions than occurred just a few thousand years before and after the LGM. Isaar et al. (1988) report that the precursor of the Dead Sea (Lake Lisan) reached its maximum depth just before the LGM, tending to indicate moister conditions. However, Bar-Yosef (1990) states that the level of Lake Lisan was actually lower than at present around 18,000 years ago, presumably indicating a switch towards a drier climate. In any case, in a tectonically active area such as this, it is perhaps questionable that the level of a single lake can be regarded as a good palaeoclimatic indicator (E. Lioubimtseva pers. comm. May 1993), and the dating of the lake phases is also an area of considerable disagreement amongst those who have worked on this area (Street-Perrott, pers. comm. 1995).
Arabian Peninsula. There are few direct indications are that Arabia was also generally drier than today, but by extrapolation from the Saharan aridity to the west and indications of lower rainfall to the south and east, it is reasonable to assume that the influence of the monsoon did not reach as far north as today. This is suggested for example by Petit-Maire et al. (1994). Large scale dune activity was more extensive than at present (Sarnthein 1982, Garcner 1988), and export of dust to the Arabian Sea seems to have occurred during the LGM period (although greater wind strengths may have contributed to this?) (Petit-Maire et al. 1994).
In fact the region may have reached its strongest aridity just after the LGM at about 17,000 14C years ago, when Garcner (1988) reports that sedimentation on the southern plains stopped completely. McClure (1985) reports lacustrine deposition in the Rub al'Hali up to 17,600 yr. B.P., and Lioubimtseva (pers. comm. June 1994) notes that a 1987 paper by Egdeff similarly reports that the aridity maximum was somewhat shifted in time from 18,000 years ago. Howeverever, the evidence in general also points to arid conditions at the LGM, and on this basis, the whole region is shown as desert on the map presented here.
Northern Africa
An extended Sahara. Desert conditions occupied a greater area to the south than at present. The southern desert boundary lay some 5 or more degrees further south than at present, with a general southwards compression of the semi-desert, scrub and woodland belts (with the sahelian scrub belt possibly disappearing). This conclusion is well established, based on a wide range of pollen, charcoal, geomorphological, zoological and anthropological evidence (see below). For instance, one of the best-known sources of evidence is the southward LGM extension of large areas of sand dunes into the present Sahelian zone (Thorp 1994 p.258). Lake level evidence from across the Sahara also agrees with the general picture of more arid than present conditions (Servant & Servant-Vildary 1989, Gasse et al. 1990, Roberts 1990).
In the uplands to the south-east of the Sahara, conditions were also drier and cooler. The Nile River, which originates in these areas, seems to have had a much lower discharge than today, for during the last glacial huge amounts of aggradational material were deposited along 1000 km of the middle Nile. These deposits contain large amounts of fresh feldspars and other unstable minerals, interpreted as the product of flash-flooding under an 'almost totally arid' climate (Fairbridge 1964) in the source areas of the Nile (including the Ethiopean Plateau?). However, there have since been discordant views on the palaeoenvironmental significance of this indicator (Adamson et al. 1980).
Temperatures in the Nile region were significantly lower than today's; for example from the distribution of 'upland' freshwater molluscs summer temperatures in Upper Egypt seem to have been about 6-7°C below modern (Close & Wendorf 1990). On the basis of a conservatively estimated temperature lowering of at least 5.5°C for mapping purposes, the 'temperate desert' boundary is here moved equatorwards and covers a large part of the Saharan region in the place of the present tropical desert.
Published map sources for the LGM Sahara. The first maps of LGM desert extent in the Sahara were put forward on the basis of dune evidence, by Grove and by Talbot. Since then, a great deal of further information has come forward. Lézine (1989) and Hooghiemstra et al. (1992) have published very useful sources indicating the position of the vegetation zones of the Sahara at the LGM and during the Holocene, based on palynological and charcoal evidence from the eastern and western Sahara respectively. As map compilations already exist for the vast amount of evidence for climate and vegetation change over most of the Sahara, little attempt is made here to repeat this work from scratch. Instead, we have based the vegetation map on pre-existing summary map sources, such as the published maps and descriptions given by Lézine and Hooghiemstra, cross-checked with each of them personally.
The LGM map presented here is also strongly influenced by the recently produced map reconstructions of N. Petit-Maire (LGQ, Luminy). She has carried out a great deal of work in compiling data on the Sahara region from published and unpublished sources, building on and improving upon the earlier syntheses of Faure, Grove and others. Her published maps of the palaeovegetation of the Sahara are based on a combination of directly and correlatively dated pollen, macrofossil, zoological, sedimentological and archaeological evidence. Lioubimtseva (1995) has also been compiling a wide range of data on these and also geomorphological and palaeosol sources, in order to reconstruct detailed maps of landscape types for the whole region for the LGM and Holocene. The landscape types in her classification represent complexes of vegetational, erosional and geological features. The results of her work give a similar picture to that of Petit-Maire; an extended Sahara in the south, where rainfall was around 150-250 mm lower than at present, but a moister area in the north-west near to the Atlas Mountains.
Moister conditions in the north-west Sahara. As mentioned above, in the extreme north-west corner of the Sahara, offshore pollen and lake-level evidence show that conditions were somewhat moister than now, with an area of mediterranean-type steppe-like vegetation having been present (Hooghiemstra et al. 1992). This steppe-like vegetation is described as being rich in Artemisia, Chenopodiaceae and Gramineae but also to some extent wooded with Pinus (Hooghiemstra et al. 1992).
In the Atlas and coastal region, well-developed palaeosols dated to around 20,000 radiocarbon years ago (just before the LGM) occur at several sites between latitudes 32 and 36N in the currently semi-arid climates of Tunisia and Morocco. Pollen spectra from these areas indicate a considerable woodland element, with around 20-30% arboreal pollen (Close & Wendorf 1990). Locally forested areas consisting of humid temperate trees may have persisted in the Atlas Mountains, which possibly had a moister climate than at present (though with lowered treelines), but these putative forests would be too small and scattered to be included in the maps here. For example, a lake site at Tigalmanine (32N, 5W) in the Atlas mountains shows a dry climate LGM vegetation with chenopods and Artemisia, but also traces of evergreen Quercus pollen (Lamb et al. 1989, Street-Perrott & Perrott 1994).
Summer temperatures in the Atlas and coastal area seem to have been at least 3-4°C lower than today, with a possible lowering of vegetation zones by more than 1200m, judging by the presence of macrofossils of indicator species of Pinus and the presence of Cedrus close to sea level (Close & Wendorf 1990). However, Street-Perrott (pers. comm. 1995) notes that such macrofossils might have been transported down from higher altitudes as they are found in large wadi systems that drain the Atlas.
There have been claims of full-glacial tree pollen and macrofossils in the Hoggar Mountains of the central Sahara (23 N), where some lakes existed during the last glacial phase. However, these are ambiguously dated and the pollen may in any case have blown in from elsewhere by long-distance transport (Tallis 1990, Livingstone 1993). Even if such trees did exist in Hoggar at the LGM, they were not at all abundant. Messerli (1980, cited by El-Nakhal 1994) reports that on the basis of periglacial features, winter temperatures in the Saharan mountains were 10-14°C colder than today.
Central and western Africa.
Plant fossil evidence of rainforest retreat. In the present rainforest region of central and western Africa, the forest area was certainly reduced to some extent relative to the Holocene, presumably as the result of aridity. One might expect a southwards compression of the rainforest zone, given the general southwards movement of the north African vegetation zones at this time. However, there is also plenty of more directly relevant evidence from around this region. At Lake Bosumtwi in Ghana, a site presently surrounded by west African rainforest, there is almost no tree pollen from the LGM period; instead grasses dominate, presenting a pollen spectrum more typical of the present Sahelian zone, but also with montane-type elements indicating at least a 3-4°C cooling (Maley 1988). By some pollen indicators the cooling would be 6°C, which is also more consistent with evidence from east Africa (Maley 1988). Livingstone suggests a no-analogue wooded grassland vegetation for this area at around the LGM, and on the abundant (>40%) presence of montane grass cuticles, indicating a cooling of at least 6.7°C. However, Maley (pers. comm. June 1994) is of the opinion that a 3-4°C cooling is more likely. Livingstone notes that the community was composed of C3 taxa , in contrast to the mainly C4 species in the area nowadays (although recent work by Talbot & Johannessen on the 13C record in the lake may suggest otherwise; Street-Perrott pers. comm. 1995)
From a pollen core situated offshore near west Africa, Dupont (1994) similarly suggests a dramatic retreat of rainforest away from the Atlantic coast in west Africa, with only tiny rainforest remnants persisting near the coast at the LGM. Further eastwards along the west African coast, there was a considerable reduction in mangrove vegetation at the LGM, but Lézine & Vernaud-Grazzini (1994) suggest that this is not due to a reduction in freshwater flux from the rivers, and that instead it relates to changes in sea level (although they do not explain in their paper the reasons for reaching this conclusion).
Grass phytoliths and charcoal carried down into the delta sediments of the Congo River may indicate that grassland-savanna vegetation was considerably more widespread in central Africa than it is at present (Caratini & Giresse 1979). In these delta sediments the percentage of pollen from forest species was very low at this time, perhaps indicating that there was very little forest vegetation within the Congo watershed. However, the actual source of this plant material remains open to question (H. Elenga, pers. comm. May 1993). It is possible that the material might have been have been carried from areas outside the central Congo Basin, perhaps from areas to the south. However, the fact also remains that there is virtually no mangrove pollen in delta sediments of this age (perhaps suggesting a considerable decrease in rainfall and freshwater discharge from the river). Also note the oxygen isotope evidence of disappearance of the Niger River flow at the LGM (see below).
Another offshore core (Jahns 1995) (core M 6-6) at 6 S, 10 E, shows a major reduction in forest pollen during glacial periods including the most recent one. Levels of forest taxa decline to less than 1/5th of those present during interglacials, but these taxa were never totally absent.
An on-land pollen core from the central Congo Basin at Bokuma-Isoku in Zaire (0 N, 18.5 E), radiocarbon dated to just after the LGM, shows what appears to be a wooded savanna environment with evidence of swamps and gallery forest along the major river courses (Preuss 1992). A core at Imbonga (0.5 S, 19 E), radiocarbon dated to just before the LGM, shows a similar picture (Preuss 1992), just before the start of sand deposition.
Maley (1987) cites another piece of indirect evidence that would lead one to believe that conditions were probably drier than present in central Africa at the LGM. He points out that LGM surface water temperatures were 2-5°C lower in the Gulf of Guinea. As lower surface water temperatures are highly correlated in the present-day with drought episodes over west-central Africa, this might have led to a similar reduction in convective rainfall, through high stratus cloud formation (Maley 1987).
Animal fossil evidence of rainforest retreat. There appears to be no evidence for the existence of an extensive LGM forest refugium at the eastern rim of the Congo Valley (presently an area of high rainfall), as had earlier been proposed on the basis of avian richness patterns; a taphonometric and palaeozoological study conducted on cave deposits in this area at 1450m a.s.l. gives no indications of any sort of forest vegetation persisting in this area in the period dated to around 20,000 14C years ago (Van neer 1984). Instead, it was concluded that the cave was located in a 'savanna environment with riverine or gallery forest nearby'.
Sedimentological evidence of rainforest retreat. There is a wide range of evidence that conditions were much drier than present in the Congo Basin and west Africa during the LGM period.
West Africa. Giresse et al. (1982) (discussed further by Preuss 1990) in a core taken at the mouth of the Niger River, found oxygen isotope evidence of a great reduction in water flow down through the Niger River for the period around the LGM. This was presumably at least partly due to a much drier climate in the rainforest areas through which it passes. In southern Ghana (present rainfall 1750-2000m) the Birim River (6 N) shows a range of features and sedimentary deposits suggesting dry conditions and very little vegetation at the time they were formed, before 13,000 BP (Thomas 1994 p.232).
Close to the Atlantic coast of west Africa, in Sierra Leone, several stream catchments which today receive >2000mm rainfall (enough to support rainforest) stopped depositing sediments in an interval radiocarbon dated to between 20,000 and 12,500 years ago (work by Thomas & Thorp, cited by Thomas 1994 p.221). This seems to argue strongly for a major rainforest retreat from Atlantic west Africa; Thomas (1994 p.221) suggests that the LGM climate at the Sierra Leone site would have been too dry even for savanna vegetation.
Central Africa. <
Quite extensive studies of stony layers and other geomorphological/sedimentological features by Runge, strongly suggest that savanna-open woodland type climates (too dry to support forest vegetation) were predominant in the eastern Congo Basin (in Eastern Zaire, Kivu-Province and Burundi) between 30,000 and 11,000 14C y.a. (Runge 1992, 1996, 1997). He suggests that the annual rainfall in the eastern Congo Basin around the LGM 1000mm, with a seasonal climate with rainy and dry seasons and high potential for erosional activity; forests would have been confined mainly to strips of 'gallery forest' along rivers. Compared to the present-day rainfall of 2000-2400mm this means a reduction by about 1000-1400mm for the LGM (not allowing for temperature lowering). Since the eastern Congo Basin at present has some of the moistest climates in the region, it may suggest that other parts of the Basin would have been even more severely affected by forest loss.
At the mouth of the Congo River, Giresse et al. (1982, 1990) find that sedimentation rate from the glacial portion (i.e. before about 12,000 years ago) of their core C237 (starting at 15,000 years ago) had a sedimentation rate some 4-5 times higher than the Holocene, with almost a complete lack of the woody material that is present throughout the Holocene portion of the core. Thus it seems that the glacial phase in the Congo Basin was characterised by erosion due to sparse vegetation and a lack of forest cover.
At the LGM, the Congo River was also transporting high ratios of well-crystallised smectite clay minerals, an indicator of contrasting seasons with a pronounced dry season (Jansen 1990). Between about 23,000 years ago and 17,000 years ago, sands were deposited over considerable stretches of the middle and lower Congo river, from about 19.5 E westwards (Preuss 1990). This phase is well constrained by radiocarbon dates from the layers immediately above and below, and appears to have reached its maximum intensity between about 18,300 and 17,000 14C years ago. The generally accepted interpretation appears to be that the sands were deposited in a braided Congo River with considerably reduced water flow, presumably due to a drier climate (Preuss 1990). Close to the mouth of the Congo, large areas of windblown dunes may have been present, for the nature of the quartz grains in the sands indicates aeolian deposition (Giresse & Le Ribault 1990).
Further north-westwards, for the West Cameroon Highlands and the nearby Adamoua Plateau, there are surface wash deposits - dated stratigraphically - of angular gravels, thought to indicate surface wash in a seasonal savanna-type environment (Tamura 1990). These areas are presently under the climate of a rainforest zone (and was forested, before anthropogenic deforestation), and this area is fairly close to the LGM rainforest refuge found in Cameroon by Maley. This seems to indicate an LGM loss of rainforest from this particular area, although the location is presently within a very steep rainfall gradient, so that not very much spatial movement of the forest boundary would be required. Kadomura & Hori (1990) have recorded carefully dated sedimentological (stone line) evidence of open grassland vegetation existing during the period 20,000-12,000 14C y.a. in southern Cameroon, an area which presently has a humid rain forest climate.
Persistence of forest and forest taxa. As mentioned above, there are indications of persistence of at least some rainforest elements close to sites which were predominantly savanna or grassland at the LGM. One definite LGM rainforest refuge has been found in West Cameroon, in a lowland (300m) site 30 km north of Mt. Cameroon (Lake Barombi Mbo), in a very high rainfall area. The cores show many lowland rainforest genera persisting through the LGM, but with tree pollen only at about 40-60% of the total, its place being taken by savanna and grassland elements such as Gramineae (in fact, the lowest percentage of arboreal pollen seems to have been reached at about 15,000 14C years ago). This indicates that unlike its present and Holocene state, the forest at this site was of an open character, with many areas of savanna. Various montane elements (e.g. Podocarpus, Olea) are attributed to cooling (Maley 1989, Livingstone 1993).
A pollen core situated south off the coast of west Africa indicates that (following an even moister-than-present phase around 22,000 14C years ago) there was a considerable rainforest element persisting near the coast at about the LGM (Lézine & Vernaud-Grazzini 1994). This seems to have been rather less than that which was present just before late Holocene deforestation. Swamp forest elements also indicate year-round river flow into the Gulf of Guinea. This conclusion of substantial persistence of rainforest through the LGM is contradicted to some extent by the geomorphological and palynological evidence of loss of forest vegetation at several sites across west Africa (cited above). Possibly, the LGM rainforest pollen flux was derived from more constricted areas of forest persisting in valleys and along rivers (Maley pers. comm.), an issue which needs further examination.
Elenga H., Schwartz D. & Vincens A. (manuscript submitted for publication, 1993) have pollen evidence from cores taken in lake basins in the Bateke Plateaux (600m a.s.l.), south-western Congo (5S, 15E), in a region of dry, sandy soils presently occupied by a rainforest/savanna mosaic. At around the LGM, conditions seem to have been drier, with an expansion of open wooded scrub vegetation at the expense of forest, relative to most of the Holocene. However, rainforest taxa did remain present in the area at fairly low levels of abundance, perhaps as gallery forest along rivers. The dry soil conditions at this site may have been especially unfavourable to forest under LGM, so perhaps forest retreat elsewhere on better-drained soils might have been less (e.g. as Dupont suggests; see below). Considerably cooler conditions are indicated by the abundant presence of montane taxa such as Podocarpus and Olea; the cooling relative to the present suggested as having been between 4 and 5°C. At the Bateke Plateaux site, Bengo & Maley find that around 40% of the LGM flora was made up of montane species such as Podocarpus and Olea, suggesting at least a 3-4°C lower temperature (Maley 1994, submitted MS).
Dupont (1995) suggests on the basis of (?new) pollen cores in the Gulf of Guinea that Podocarpus forest rather than savanna was widespread through west Africa and the Congo Basin during the LGM. However, Maley (pers. comm. August 1995) is in disagreement with this view, pointing to the broad range of sedimentological evidence which indicates loss of forest cover at the LGM. Lézine (pers. comm. August 1995) is also in disagreement with Dupont's reconstruction, suggesting that the complexity of winds and surface currents in the Gulf of Guinea does not allow realistic mapping of the vegetation that was the source of the pollen. Both Maley and Lézine instead suggest a widespread forest reduction in central and western Africa.
Forest refugia based on present-day biogeography. On biogeographical grounds (usually species richness patterns, or the positions of boundaries between the ranges of closely related species), various forest refugia for the rainforest region of central Africa have been suggested, although these must remain tentative in the absence of direct evidence. It is important to bear in mind that an LGM refugial area was not necessarily extensively forested; all that is necessary is the presence of some forest (e.g. along rivers), although no-one can be sure just how much forest would be needed for all these species to have survived.
An LGM forest refuge area (or set of smaller refuges) has been suggested for coastal SW Gabon in a doctoral thesis by M.S.M. Sosef, on the basis of present-day diversity patterns of Begonia spp., the theory being that these are very poorly dispersing species which would take many thousands of years to expand its populations out from refugial areas (Sosef 1991, Maley 1994, submitted MS) (note that this area was also predicted as a possible rain forest refuge by Maley 1987, on the basis of present-day rainfall patterns). Sosef notes that endemic species also occur in Cameroon, in the same area where palynological evidence indicates forest survival.
For the Congo Basin, various sources of biogeographical evidence have been taken as indicating or confirming one forest refuge area in south Cameroon, another along the rivers of the central Basin, and a third in Gabon. The distribution of human pygmy tribal groups has also been suggested as evidence by S. Bahuchet (cited by Maley 1994, submitted MS). An east-central refuge of the African rainforest has also been suggested by Hamilton (1982) on biogeographical grounds (present-day patterns in bird species richness and endemism) for the elevated eastern extreme of the central rainforest block, in eastern Zaire, but palaeozoological and sedimentological evidence from the heart of this proposed refuge (see above) seems to argue strongly against this possibility. The idea of an east-central rainforest refugium at the LGM has also been attacked on biogeographical grounds by Colyn et al. (1991), who suggest that a central Congo forest refugium is more likely, on the basis of present-day monkey distributions.
Bengo & Maley (1991) point out that the floristic differences between the Zambezian (southern) and Sudanean (northern) savannas suggests that they have never been in direct contact across the equator. This may indicate a montane-type forest or woodland barrier existing in the Congo Basin during cold periods, although the authors (pers. comm.) appear to favour the hypothesis that there was for the most part an open vegetation such as the open savanna which currently grows on the Bateke Plateau to the south-west. Maley suggests (pers. comm. Sept. 1991) that at least some gallery or flood plain forest has always persisted in Central Africa along the Congo river during the last million years or so, helping to provide the necessary barrier between the plant taxa of northern and southern savannas.
Present-day biogeographical evidence of lower temperatures. The presence of large numbers of species of mid-altitude grasses and birds in common between Mt. Cameroon and the east African mountains suggests an extreme cooling of perhaps 10°C, for them to have spread directly across the intervening lowlands at some time in the past - though not necessarily at the last glacial or the LGM itself (Livingstone 1993). Maley (pers. comm. May 1994) is sceptical of Livingstone's interpretation of such a large temperature lowering, pointing out that more direct indicators do not show such a large change. However, perhaps a less complete extension around part of the raised rim of the Congo basin might have been enough to allow this transfer? (pers. obsv.n). Note that in contrast to this mid-altitude east-west link, higher montane species do not show this resemblance, suggesting that temperatures have never been low enough (about 16°C lower) to allow these species to spread across the lowlands.
Overall; a substantial forest reduction. There can be no doubt that the area of rainforest in central and western Africa was reduced at the LGM, but the question is by how much. The plant fossil evidence points to a considerable LGM forest reduction in at least some specific areas, as do the more indirect clues from animal fossils, biogeography and in particular sedimentology. The combined evidence seems to strongly suggest a major reduction in forest cover during the last glacial, but most probably at least one substantial area of forest surviving in the low-lying central Congo Basin, and other smaller areas in west Africa and (with savanna openings) in Cameroon. This general pattern of reduced forest cover also agrees with that found on all sides in other areas of Africa, where greater aridity is found for the LGM.
Livingstone (1993) concludes that as yet it is impossible to know whether the rainforest that turns up in Cameroon at the LGM was confined to a very small area, or whether it extended as an almost continuous cover south to Gabon or even through the Zairian cuvette to the moist highlands in the east. This seems an unduly agnostic view, in the light of all the fossil and geomorphological/sedimentological evidence for a considerable loss of forest from various parts of the Congo Basin. In our reconstruction here, we suggest that the various sources of evidence taken together suggest a strong reduction in rainforest area for the LGM, but with substantial areas of forest remaining along the low-lying floodplains of the major river courses, in areas currently occupied mainly by swamp forest.
Aridity and reduction in forest cover was not the only factor worth mentioning, however. The combination of microfossils and present biogeographic ranges tends to suggest temperatures in the lowland rainforest zone around the LGM that were at least 3-4°C and perhaps 6°C lower than today. This fits in with the picture emerging elsewhere in the moist tropics for the LGM.
East Africa and the Horn of Africa. In east Africa, the general picture for the LGM is one of drier-than-present conditions throughout (Coetzee & van Zinderen Bakker 1988). Climates were also substantially cooler, according to pollen evidence and from the lowering of glacier limits by about 1000m.
The east coast. Hamilton (1982, 1988) suggested a substantial refugial area of woodland and forest, in the form of coastal forest along the east coast of Africa. This was on the basis of biogeographical evidence, but there seems to be no pollen evidence to support the existence such an extensive band of forest or woodland. Extrapolating from the regional trend towards drier conditions, we suggest that this belt would have been much reduced or fragmented.
Drier conditions throughout the rift valley region. The general picture throughout this region, from the southern Ethiopean rift lakes to the western/central rift lakes (e.g. L. Kivu, L. Victoria), and L. Malawi in the southern rift, is of drier conditions before 13,000 14C years ago, although not all of the lake level records go back as far as the LGM (Hamilton 1982). Lack of pollen evidence makes it difficult to guess what the vegetation in some areas (especially Ethiopea) would have been like, although it would presumably have been sparser with less of a woody component.
Disappearance of the montane forests. At present, there are considerable areas of moist montane forest throughout the rift valley uplands, at altitudes between about 1500 and 3000m. Hamilton (1982) noted the persistence of montane forest during the LGM at Kamarizovu in the Ugandan Highlands. Yet evidence from several other sites (Lake Rutundu, Mahoma Lake, Sacred Lake and Muchoya Swamp, on Mt. Kenya, Ruwenzori Range, Cheragani and Mt. Kenya respectively) shows that at the LGM this forest was completely lacking, presumably reduced by drought and by lower temperatures (pollen evidence reviewed in Street-Perrott & Perrott 1993, and Flenley pers. comm., D. Olago pers. comm. Dec. 1994). Only a very sparse woody component seems to have been present locally at Muchoya Swamp and Kashiru bog (see below).
Pollen evidence (Street-Perrott & Perrott 1993) shows that the present-day forests were replaced by sparse, dry afroalpine vegetation (a dry tundra-like vegetation on rocky soils) above 2000m. Below this there was a belt of dry ericaceous vegetation, where there were some trees present locally, and below this was a dry open grassland vegetation. Thus for instance at 2350m on nearby Mount Kenya, at Sacred Lake, there was the dry ericaceous vegetation in the place of the present moist montane forest (Coetzee 1967, Street-Perrott 1994). It seems that conditions at this altitude were both cooler (by between 8.8 and 5.1°C) and considerably drier.
In the highlands of Burundi, pollen influx indicates a loss of the present forest cover, but with some woody components remaining present in the landscape, under generally somewhat cooler and drier conditions than at present (Bonnefille & Riollet 1988). Aucour et al. (1994) suggest that the dominance of C4 plant species in a highland peatbog (Kashiru) in Burundi during the LGM was due to lower atmospheric CO2 levels favouring these plants over the less CO2-efficient C3 species. These are under conditions (lower temperatures, but apparently almost constant local water table conditions relative to the interglacial) that would instead have been expected to favour a shift towards C3 plants relative to the Holocene (R. Bonnefille pers. comm.).
On the basis of such evidence, we conclude that the LGM upland forest cover in east Africa was too small to map, on the scale at which these maps are drawn.
Lowlands of the central rift valley; drier conditions, drier grasslands. Pollen from a lowland lake in the central rift valley region, west of Mt. Kenya, 1 S, 37 E (Maitima 1991) indicates drier conditions with replacement of the present woodland and savanna-woodland by grassland in the lowlands around the equatorial part of the rift valley (Ssemmada & Vincens 1993). Street-Perrott (1994) presents a sketch diagram (originally from the PhD thesis of D. Olago) of the change in vegetation distribution on Mt. Kenya, based on these various sources of evidence, showing much less woody vegetation.
The picture of drier conditions is reinforced by lake level evidence. The plains south of mount Kenya supported populations of arid-adapted ungulates such as oryx, suggesting arid grassland conditions instead of the treed grassland that exists there today (Marean 1992).
A similar picture comes from the Cherangani Hills, just to the north of Mount Kenya (van Zinderen Bakker & Coetzee 1988), and also from the mountains to the west of Lake Victoria. From his data at Lake Mobutu in Uganda, Harvey (1976, PhD thesis cited by Livingstone 1993) suggested LGM rainfall to have been about 30% lower than at present, though details on what the corresponding vegetation type might have been are not given in this citation.
Southern rift valley ; more open vegetation. In the more southerly part of the rift-valley region (approx. 3 - 9S), in the Lake Tanganyika basin, 14C-dated pollen cores indicate that instead of the present woodland vegetation there was a more open (savanna-like) woody cover in the lowlands, with local development of a mixture with montane forest patches on highland areas at above 1500m, and fragmentation of the montane forest that now occurs above 2000m (Vincens 1991). A recent study of lake level evidence of Lake Tanganyika also indicates a peak of aridity, relative to the present, at around 18,000 years ago (with the lake reaching its general interglacial level by around 12,700 years ago) (Gasse et al. 1990).
An onset of stream channel downcutting in Malawi and Zimbabwe after around 12,000 years ago may also indicate a shift towards moister climates in the Holocene following glacial-age aridity, although this view is open to dispute (Thomas 1994 p.278).
Ethiopean Plateau; more arid? Based on biogeographical evidence, many reconstructions assume the persistence of large areas of woodland and forest vegetation in the highlands of the Ethiopean Plateau in the north-west, although there seems to be no direct evidence for a large woody cover persisting. One indicator of general climate conditions on the Ethiopean Plateau is the adjacent lowland Lake Abhe (11N, 42E, 240m a.s.l.), which receives most of its water from the Plateau via the Awash River. From the radiocarbon dates and the stratigraphic diagram given by Gasse (1975), it seems that the peak of aridity - when the lake dried up completely - was not reached until after 17,100 years ago (the lake also stayed dry up until 10,000 years ago). From Gasse's dating evidence it seems possible that 18,000 years ago was also rather more arid on the Plateau than today, though presumably not as arid as after 17,000 years ago. There is evidence from the Arabian region (Adamson et al. 1980) to the north of a similar aridity peak around 17,000 years ago. Lake Turkana, which is fed from the Ethiopean Plateau, was also lower than at present during the LGM period, again suggesting lower rainfall in its highland source areas.
As mentioned above, there are strong indications of drier conditions in the two main Nile headwater regions during the last glacial period. Analysis of the changing trace-element composition and particle size of the sediments of the Nile is seen as reflecting changes in the relative contributions of the central African (White Nile) and Ethiopean uplands (Blue Nile), and may point to a sparser vegetation cover in the Ethiopean Plateau during the LGM (S. Stokes, School of Geography, Oxford University; seminar, January 1994; also results partially published in Allan et al. 1992).
Conditions over other parts of Ethiopea are a matter of conjecture. With the evidence of drying in the highlands and the rift valley region, we suggest that tropical desert covered much of the Horn of Africa.
South-Central Africa.
Drier conditions? There seems to be very little evidence of last-glacial environments from the area stretching across from Angola and Zaire to Mozambique. Fairbridge (1964) notes that the Zambezi River during the last glacial period shows a similar siltation phase to that observed for the Nile and other Saharan/Sahelian rivers during the same time span, indicating arid conditions with occasional flash floods from sparsely vegetated landscapes in its headwaters. Thomas (1994 p.258-9) also refers in passing to the occurrence of fans and dunes associated with shallow clay and salt pans, close to the present Zambezi headwater regions in the central African plateau in south-eastern Angola and western Zambia. These areas may have been formed under arid internal drainage conditions, and Thomas suggests that the pans were formed subsequent to a drier phase during which the dunes were formed (see below). In any case, the area now has a rainfall of about 600-800mm, with scrub and woodland vegetation, and these features are presently dissected by stream courses. The evidence of extensive LGM dune activity from the last glacial just south of the Zambezi (Stokes et al. 1997) would seem to reinforce the view that this could be part of an LGM arid phase across the region (see below).
A core obtained by Livingstone (1971) from the Zambezi region dating back to 22,000 14C years ago, indicates a fairly similar LGM pollen spectrum to that existing at present. However, because the spectrum is dominated by leguminous species, which cannot easily be separated by their pollen types, it is not possible to constrain the true character of the climate or vegetation cover during this time period. For example, variations of +/- 50% in rainfall could have occurred without any detectable effect on the composition of the pollen spectrum (Livingstone 1971).
Southern Africa.
Wetter conditions? Brook et al. (1987) note that various lakes in south-central Africa suggest moister-than-present conditions close to the LGM or during the late glacial (possibly due to either increased summer or winter rainfall). These include sites in northern Cape Province, NW Orange Free State (South Africa), eastern Namibia, Tsodilo Hills in Botswna (Kalahari Desert) and Paleo Makgadikgadi Lake in Botswana. A.T. Grove (pers. comm., May 1996) is somewhat sceptical of the validity of these dates because the lake sediment calcite from which the dates were obtained is likely to contain 'ancient' carbon that would give a misleadingly old date for a lake phase. There is contradictory evidence from across the region, in the form of palaeobotanical, dune and zoological evidence, which suggests that in fact the LGM period was indeed drier than at present. Recent dates on the extensive dune activity in northern Botswana (Stokes et al. 1997) seem to confirm that most of the area was indeed very arid (see below).
The western part; mainly desert? The Namib desert seems to have been even drier than present at the LGM, and was possibly expanded in area (Deacon & Lancaster 1988), both eastwards and northwards. Northwards expansion is indicated by the high ratio of illite clays and plant phytoliths turning up in the glacial-age sediments off the coast of northern Angola (Jansen 1990). In the Namib, lake and groundwater levels fell and the driest conditions occurred at around 19,000-18,000 14C years ago. Sarnthein's (1978) map shows active dune fields covering most of the area of south-west Africa. This would seem to indicate very sparse vegetation throughout this area. There are at least some dates available; for the Kalahari sands for example (20S), Sarnthein (pers. comm., 1994) refers to radiocarbon evidence from papers by Grove and by Cooke for the dunes becoming stabilised about 17,300 yr. ago. The contradictory view that conditions might have been moister than present in parts of the Kalahari has now been abandoned by at least some who work in the area (e.g. T. Partridge, Transvaal Museum pers. comm. June '93) on the basis of dating evidence that confirms at least parts of Sarnthein's picture of dune fields in the region at the LGM. Some recent data and interpretations are summarised in Thomas & Shaw (1991). in northern Botswana (19-17 deg.S) conditions were much more arid than present with extensive dune activity suggesting widespread desert conditions across southern Africa (Stokes et al. 1997).
Partridge (1997) has reconstructed LGM rainfall isohyets over southern Africa for the LGM, on the basis of many different sources of evidence combined. He suggests that in sw Zambia, SE Angola (about 15 deg.S) was about 20% of present values (about 150-200mm compared to the present 900mm or so), with extreme desert and extensive dune mobility. Most of SW Africa seems to have had rainfall about 50% of present values, enough to give widespread desert. The SE of the region is suggested as having higher values, about 60-70% of present-day. It is not clear whether these reconstructed values take account of temperature change.
Further east, dry and cool conditions. Over this region as a whole, the picture for the LGM is of drier and cooler conditions (about 5 - 7.5°C cooler in the lowlands) based on pollen, speleothem isotopes and zoological evidence (Walker 1990). There was much more open scrub and sparse grassland, in areas that now support a dense woody vegetation or steppe (Walker 1990).
At the Pretoria Saltpan in the north-eastern part of South Africa (about 26 deg.S, 28 deg.E), precipitation (not allowing for temperature changes) inferred on the basis of sediment grade seems to have been about 30% lower than present at the LGM (about 560mm as compared to the present 660mm) (Partridge 1997). This can be expected to have given a greater proportion of dry grassland vegetation. For south-eastern Africa in general, Partridge (1997) suggests that the effective rainfall was about 60-70% of present-day values.
Dry conditions in the Cape. In the SW Cape winter rainfall area, conditions seem from zoological evidence to have been rather drier than at present (Deacon 1987). However, Cape heathlands and dense scrub may have persisted in the Cape fold mountains in places where the soil favoured this, as it is a vegetation type whose distribution is controlled more by soil than by climate (Deacon 1987).
Semi-desert over most of the central and eastern areas? Over most of central and eastern South Africa (where shrubby karoo or woodland vegetation now predominates), a drier semi-desert karroid scrub seems to have been present over most of the area, based on various pollen sites (Deacon & Lancaster 1988, Deacon 1990). Lancaster (pers. comm. Jan. 1990) suggests that there may also have been arid semi-desert grassland in what is presently the more densely vegetated southern Kalahari.
Dry grasslands in the south-east. Towards the south coast, pollen and charcoal analyses at Boomplas show a no-modern-analogue vegetation which had few trees and a high percentage of compositae, in an area that today has woodland vegetation. Zoological evidence here, and also by the present coastline to the south, also agrees in indicating much drier-than-present LGM conditions (Deacon 1990). J. Deacon (pers. comm. Feb. 1990) suggests on the basis of coastal faunal remains that the clayey substrates would have favoured grassy vegetation on the exposed shelf area to the south. Van Zinderen Bakker and Coetzee (1988) also suggest grassland along the coast in the south-east.
Dry conditions and lowering of vegetation belts in north-east. Likewise towards the north-east of South Africa, in the Orange Free State and the Transvaal, pollen evidence from sites scattered through these areas generally shows impoverished vegetation indicative of relatively dry conditions at the LGM (Scott 1989, Deacon 1990). However, denser 'bushveld' scrub may have survived in parts of the uplands to the north-east, especially north of the Limpopo River (Deacon pers comm. Feb. 1990, Cowling pers. comm. Feb. 1990). At Wonderkrater (24 S, 28 E) in the Transvaal (Scott 1982), the pollen spectrum before 14,100 14C years ago indicates grassland vegetation under a cool to cold-temperate subhumid climate (moist but cooler than at present), with traces of distant montane forest pollen. The cooler climate would have brought down upland vegetation belts by around 1000m, relative to their present positions. A pollen-bearing core at Elim indicates that the highveld grassland of the Transvaal (which presently occurs above 1600m) may have been largely replaced by the upland grassland that presently occurs above 2100m, but with a strong fynbos element in this LGM grassland indicating drier-than-present conditions (Scott 1989). Partridge (pers. comm., June 1993) notes that his evidence from the Pretoria Saltpan suggests an aridity maximum about 4,000-5,000 years after the temperature minimum, though he does not state when this is.
Madagascar. Drier and colder. There is very little evidence of Quaternary palaeovegetation from the island of Madagascar. Published work by Straka (pers. comm. A.A. Velichko, April '92) apparently indicates drier-than-present LGM conditions. Sarnthein (1978) shows active dunes in south-western Madagascar on his LGM map, but the source of this information and the nature of its dating are not clear from the text. The only reference to glacial-age dunes that we have found comes from a report of 'pre-Flandrian' age dune activity in southern Madagascar, occurring during an extreme low sea level phase presumably corresponding in age to the part of the last glacial period (Battistini 1970). Gasse et al. (1994), in a core from a crater lake on the Malagasy Plateau, have found evidence of much lower water tables during or about the time of the LGM, with oxidation of lake sediments. Pollen of Ericaceae is relatively abundant at this time, indicating considerably cooler conditions than at present.
8,000 14C years ago (early Holocene)
The Levant. Moister conditions with coastal forest. The vegetation maps shown in van Zeist & Bottema (1988), on the basis of two inland pollen localities, suggest slightly moister conditions than at present. A fringe of closed forest extended along the coast and about 70 km inland. Inland from this was a belt of forest steppe extending another 50 km inland, in what became treeless vegetation by the later Holocene. Most of the area further inland seems to have been semi-desert, as today. Mordeckai and Goodfriend (1987) mention various sources of evidence which they take to indicate moister-than-present conditions between 8,000 and 9,000 14C years ago. For example, in the Jordan valley there was increased human habitation and farming in areas which are now too dry to farm. In Syria, high levels of arboreal pollen are recorded just before 8,000 years ago. In the Northern Negev, C3-rich vegetation characteristic of the zone some 30km to the north in this steep rainfall gradient, was present instead of the sparser C4-dominated type of semi-desert that exists at present. This zone now has <150mm annual rainfall, but at that time the rainfall must have been about 300mm. Likewise, the central Negev, which is now 'very arid' (<100mm annual rainfall) seems to have been 'not particularly arid' in the words of Mordeckai & Goodfriend (1987).
Arabian Peninsula. Given that the Sahara region (see below) was so much moister at around this time, one would expect the adjacent Arabian Peninsula to have experienced roughly similar conditions. Yet in contrast to the Sahara, ther e seem to be relatively few data available. Al-Sayari & Zoetl (1978) report that isotopically dated piedmont gravel sequences indicate a pluvial episode 9,5 00-4,500 years ago in eastern and central Saudi Arabia. Whitney (1983) confirms the same general picture of pluvial events in western Saudi Arabia. Lake deposit s dating to around 8,000 years ago are recorded, in areas where there are now no lakes present (Hotzl et al. 1984, Roberts & Wright 1993). On the presently waterless Dahalk Islands in the Red Sea, there was enough freshwater present to allow an early-to-mid Holocene freshwater mollusc assemblage to exist (radiocarbon dated to around 6,500 years ago) (Belluomini et al. 1980). Pollen evidence from lakes in southern Arabia during the lower Holocene suggests a semi-arid scrub vegetation, with the low levels of sedge pollen indicating that sand dunes were stabilised by a vegetation cover at that time (Roberts & Wright 1993). In summary, there is evidence of moister-than-present conditions in the Arabian Peninsula between around 9,000 and 6,000 14C years ago, particularly in the south and the central region, and also at least as far north as the Nefud (presently an area of sand desert, between 29-27N, 40-44E) (Petit-Maire et al. 1994). This is in accordance with the northwards shift of the monsoon belt in the Saharan region to the west, and in southern Asia to the east. Whitney et al. (1983) suggest that annual rainfall during the early Holocene was about 250-300mm over much of the Arabian Peninsula (as opposed to the present 50-100mm); enough to sustain a semi-desert rather than the present desert vegetation. Petit-Maire et al. (1994) go further to suggest that rainfall in Arabia was 100-400mm higher than now during the early Holocene, with the monsoon rains reaching hundreds of kilometres further north than at present. Lioubimtseva (1995) suggests in her review that on the basis of the combined evidence, sclerophyllous woodlands would have dominated in the highlands of western Arabia, whilst steppic landscapes were widespread on the plains of northern Arabia. On the basis of such data and interpretations, and by extrapolation from the adjacent Sahara region, we suggest a mixture of grassland and semi-desert covering the Arabian Peninsula at around 8,000 years ago.
Sahara region.
Fossil evidence for vegetation extending over the Sahara. The Sahara desert in its present sense seems to have disappeared in the early Holocene due to much higher monsoonal rainfall. Pollen-based reconstructions of vegetation zones are available from various sources for the period around 8,000 years ago. Published maps that attempt to reconstruct the vegetation zones include one for the western Sahara, reconstructed from offshore cores taken by Hooghiemstra et al. (1992) for the western Sahara, and another from charcoal evidence in the central Sahara by Lézine (1989), indicating that the southern vegetation belts lay some 400-500 km north of their present positions at 8,000 years ago. For the eastern Sahara, Neumann (1989) has reconstructed vegetation zones for the period 7,000-6,500 14C years ago, again based on charcoal evidence. She finds that at this time, the south Saharan vegetation belts lay much further north than at present, about 500-600 km north of their present boundaries. However, it was not a case of simple shifting in latitude; to some extent savanna and sahelian scrub vegetation seem to have formed mosaics in the landscape, in a way that is not seen today (Neumann, pers. comm. Feb. 1990). Likewise, the semi-desert vegetation that exists within 100 km of the northern (Mediterranean) coast at present, seems to have extended southwards by more than 500 km, to about 26 N. Although a reconstruction for 7,000 years ago might not be representative of 8,000 years ago, we have taken this as our 'nearest guess' of what the vegetation was like in the eastern Sahara.
A pollen study by Ritchie (1994) in the eastern Sahara reinforces the existing picture, suggesting a mosaic of deciduous savanna woodland, scrub vegetation and lakes between 9,000 and 6,000 14C years ago at Oyo (19 N, 26 E) in north-eastern Sudan, an area which currently receives less than 5 mm rainfall each year. A somewhat similar savanna-type vegetation currently exists at around 12-15 N. Although Ritchie cautions against the hazards of reconstructing palaeovegetation belts on the basis of sparse pollen evidence (noting the no-present-analogue nature of the vegetation mosaic that existed at that time), he does venture to suggest that a north-south zonation resembling the present existed in the eastern Sahara, but that it was weaker and shifted far northwards in latitude.
Note that Tyrberg (pers. comm. July 1998; tommy.tyrberg@ffvaerotech.ffv.se) finds biogeographical evidence from avain fuanas that there has always been some sort of extreme desert boundary in the eastern Sahara, preventing certain sahelian forms from reaching the north coast. He notes that there is not direct evidence of moister-than-present conditions in the western desert of Egypt, and hypothesises that it always remained hyper-arid, even in the early Holocene and during the previous interglacial.
Other sources of evidence for a moist Saharan climate. This general picture of much moister-than-present conditions at around 8,000 14C years ago is corroborated by a wide range of non-palaeobotanical sources of evidence all across the Sahara. Dune activity seems to have been much reduced at that time, compared to the present (Sarnthein 1978). Lake level (Servant and Servant-Vilardy 1980), zoological, geomorphological and anthropological evidence all point to much moister conditions than at present at 8,000 years ago (N. Petit-Maire, pers. comm., 1991). Although a relatively dry episode does seem to have occurred around 8,200 years ago in the lake level record, by 8,000 years ago lake levels had recovered (but note that in west Africa, Gasse & Van Campo 1994 also find evidence for a dry phase just after 8,000 years ago). Generally it seems that the Saharan climate was at or very near its 'optimum' (i.e. wettest) phase about 8,000 years ago. As a result of an extensive review of the data on Holocene Saharan environments, Vernet (1995) suggests that the wettest and most vegetated period in the Sahara was between 8,500 and 7,500 years ago. In a recent summary of the lake level evidence, Damanti & Harrison (1995) note that the highest Holocene lake levels occurred all across Northern Africa between 9,500 and 7,000 years ago. From planktonic isotope ratios and sedimentary indictors, the Nile seems to have attained its maximum discharge at around 8,000 years ago, reflecting both rainfall in the source areas of the White and Blue Nile and also over the Sahara region itself (Thomas 1994). Likewise, Fairbridge (1964) cites evidence of greatly increased river discharge resulting in rapid downcutting by the Nile during the early Holocene. Very high freshwater discharge appears to have been the cause of a sapropel (organic mud) layer deposited in the eastern Mediterranean between 9,000 and 8,000 years ago (see evidence discussed by Thomas 1994 p.219).
Petit-Maire has compiled map evidence from the whole range of sources, palaeobotanical or otherwise, to arrive at map reconstructions of vegetation and rainfall belts during the early Holocene. There is considerable evidence that all through the Sahara belt there was abundant vegetation. For instance, Baumhauer (1995) suggests on faunal, pedological and lake-level evidence that at a central Saharan site in N.E. Niger (presently an extensive sand plain), annual rainfall during the lower Holocene (before about 6,500 years ago) was greater than 400mm, enough to support a sahelian or dry grassland vegetation. Current field research (as yet unpublished) shows that even the western desert of Egypt, which is now the driest part of this hyperarid region, had a diverse flora and mammal fauna at around 8,000 years ago (and supported abundant early crop agriculture and herding). This suggests a climate and ecology at least comparable with the present-day sahelian zone, or even moister (S. Stokes, pers. comm. Jan. 1994).
Lake Mega-Chad. Perhaps because of a greatly increased river influx from the Equatorial zone to the south (which currently provides 80% of its present water input), Lake Chad occupied several times its present area (forming 'Lake Mega-Chad') for most of the time between around 9,000 and 4,000 14C years ago (Maley 1977), and was probably surrounded by extensive Papyrus marshlands (indicated by surviving peat deposits) and a mosaic of inter-dune lakes fed by groundwater (Maley 1977). Lake level fluctuated considerably, but the approximate maximum lake extent (about 350,000 sq.km) persisted at least until after 6340+/- 250 B.P. (Thiemeyer 1995). According to Servant & Servant-Vilardy (1989), Lake Chad had a temperate rather than tropical diatom flora around 8,000 years ago, possibly due to occasional incursions of cold northern air.
In the mid-Saharan mountains (Ahaggar, Tassili-n-Ajjer & Tibesti), pollen assemblages have been taken to suggest at least some forest or woodland vegetation at 8,000 and 5,000 years ago, including temperate deciduous tree taxa such as Tilia (Schulz 1986, Street-Perrot & Perrot 1993). This view is regarded as unlikely on the basis of more recent studies by Thinon et al. (1996) of pollen-bearing sediments from the Ahaggar massif. Thinon et al. instead suggest that the flora of the Saharan mountains has remained very similar throughout the Holocene, and that the impression of temperate and mediterranean taxa is derived from long-distance transport of pollen
Northern-western Sahara/Mahgreb. Fairly similar conditions to the present. In the north-west, after 8,500 years ago there was an expansion of mediterranean oak (Quercus) woodland/forest cover between the Mediterranean coast and Atlas range. Pollen evidence from various sites in the Atlas region suggests a woody cover similar to the present-natural, although generally with increased moist climate elements (Lamb et al. 1989, Hooghiemstra pers. comm. July 1992, Street-Perrot & Perrott 1995). However, arid conditions drier than present may have existed on the Saharan Atlas slopes, and perhaps further east on the south-eastern Mediterranean coastal areas (Lioubimtseva 1995).
Central and western Africa.
Wetter, more forested conditions within the present rainforest block. By 8,000 14C years ago, the forest seems to have regained the area that it had lost during the last glacial. There are some indicators (e.g. sediment flux from the Congo River) that indicate no difference from the present or late-historical situation, in terms of vegetation cover and climate. However, there are also reasons for thinking that there would have been a greater-than present area of evergreen rather than semi-deciduous forest, existing under a higher-rainfall regime. For both 8,000 years ago and 5,000 years ago there are various indications of denser forest and higher rainfall conditions within the present rainforest region. At Lake Barombi Mbo in West Cameroon, the pollen spectrum between about 10,000 and 3,500 years ago was about 98% arboreal, after which climatic drying followed by agricultural impact appear to have set in (Maley 1992). The forest had also returned to the site in Ghana from which it was lacking at the LGM. Gasse & Van Campo (1994) have compiled lake level evidence from across from west Africa to south Asia, suggesting a dry phase after around 8,000 years ago. However, at 8,000 years ago itself, conditions would still seem to have been very moist.
Rainforest expansion at the northern edge. Central African rainforest se ems likely to have extended much further north than at present, probably filling the gap between Benin & Togo (J. Maley pers. comm., June 1994). One would certainly expect, from the much further northward penetration of monsoon rains into the Sahara at this time, that the rainforest zone would likewise have been much more extensive. The presence of rainforest pollen elements in Lake Chad also seems to indicate at least some northwards expansion of rainforest, although in fact the rainforest did not reach as far north as Chad itself; the pollen was simply transported down by rivers, and derived from gallery forests along these rivers (J. Maley pers. comm., June 1994). In a core just off the coast of west Africa, in the Gulf of Guinea, Lézine & Vernaud-Grazzini (1994) also find a considerably increased flux of rainforest pollen elements (compared to the late Holocene), centred at around 8,500 14C years ago. Likewise, there seems to have been a greatly increased freshwater flux, relative to the present, from the Niger River, suggesting higher rainfall conditions within its catchment area (Giresse et al. 1990). At the same time, there seems to have been a much greater than present extent of mangrove vegetation - perhaps an indicator of greater-than-present fluxes of freshwater down the rivers? However, from a core to the west of the northern 'present natural' rainforest boundary next to the Atlantic coast of west Africa there seems no evidence of a relative northwards expansion of forest at 9,000 years ago (Dupont 1994). On the map presented here, the 8,000 year northward limit of rainforest is shown somewhat further south than the earlier reconstructions of Hamilton (1982, 1988), on the advice of Maley (pers. comm. June 1994), but roughly corresponding to the northward movement of Saharan isohyets suggested by Petit-Maire and others. Nevertheless, the precise position of the rainforest boundary must remain a matter of some conjecture.
There is uncertainty over whether there was also a large early-to-mid Holocene rainforest expansion south of the Equator. No doubt, even if climate was much the same as today's, the forest would have been more extensive because the present-day 'natural' forest extent is almost certainly limited by a history of human disturbance (especially burning of savanna vegetation) which may not have been as severe in its effects in the early Holocene. Hamilton (1982, 1988) has in fact suggested that the maximum early-to-mid Holocene extent of rainforest in central Africa covered nearly all of the present 'forest-savanna' mosaic zone of White (1983), extending hundreds of kilometres both north and south of its present range.
There is in fact palaeoenvironmental and fossil evidence for a substantially greater southward extension of rainforest by around 8,000 years ago. From analysis of soil carbon at several sites, Schwartz (1991) has found that areas that are presently covered by savanna across the south-west of the rainforest region 0-5 S, in south-western parts of Congo, were covered by C3 plants - almost certainly forest - at around 8,000 years ago. A broadly similar picture of forest expansion and moister-than-present conditions is found by Elenga et al. (1993) from pollen evidence on the Bateke Plateau, south of the Equator. However, Elenga et al. emphasise that at least some areas of the savanna mosaic persisted throughout the Holocene without ever being totally forested in this raised area of relatively droughty sandy soils. In this study, Elenga et al. find evidence for an considerable expansion (relative to the present) of the proportion of forest vegetation in the lowland parts of what is presently the forest-savanna mosaic in the Plateau region, and also an expansion of woodland relative to savanna on the higher slopes.
East Africa.
Similar-to-present montane vegetation cover. Montane forest cover at 8,000 14C years ago seems to have been much as it is at present. The various sites (Street-Perrott & Perrott 1993) referred to above show a return of montane forest to its present limits by about 8,000 years ago (Flenley 1979), with much the same being present at 5,000 years ago. The Ericaceous vegetation may not have reached its present upper limit until shortly afterward 8,000 years ago, on Kitandara at least (Flenley 1979).
In the mountains of east Africa, pollen diagrams thus indicate vegetation conditions very much like the present at both 8,000 years ago and 5,000 years ago (Hamilton & Taylor 1991). Maitima (1991) comments that there was dry forest on the mid altitudes of the Nyanduru Mountains just to the west of Mt. Kenya at 8,000 years ago, but he does not comment on how this might have differed from present vegetation.
Moister conditions with more woody vegetation in lowlands. In East Africa, conditions at 8,000 years ago seem to have been generally moister than present, from lake level and pollen evidence. Relatively humid Zambezian types of woodland vegetation were more widespread than today along the rift valley region of Africa, south to Lake Tanganyika, including some taxa that today are more characteristic of the humid west and central African regions (Vincens 1991). In the central rift valley of Kenya, lake pollen evidence (36.5E, 1S) indicates woodland or forest vegetation throughout the area in the early-to-mid Holocene (12,200-6,500 years ago), whereas the vegetation is now largely grassland (Maitima 1993). This is interpreted as having been due to a moister climate, although increased human influence may also have altered the vegetation over the past couple of millennia.
The levels of Lake Abhe and Ziway-Shala in the Ethiopean Rift were considerably higher than at present, indicating moister conditions in the Ethiopean Plateau and in southern Ethiopea respectively (Gasse 1976, Hamilton 1982). However, the levels of Lake Kivu and Lake Mobulu, in the western rift (Uganda) and also Lake Victoria were about the same as at present, indicating a generally similar-to-present moisture balance, in contrast to the vegetation evidence of moist climates (Hamilton 1982).
Fairbridge (1964) mentions in passing that the Zambezi River shows strong downcutting following the drier aggradational conditions of the last glacial, and then a partial return to drier conditions later in the Holocene. The implication (which is not clearly specified) seems to be that conditions were moister than present in the lower Holocene, resulting in a higher river discharge.
On the basis of the indications of generally moister conditions in east Africa during the early Holocene, Hamilton has suggested that the east African fringe of coastal rain forest was expanded inland at that time. This expansion, whilst not backed up by any other evidence, is likewise tentatively suggested on the map given here.
Southern Africa. Generally similar to the present. Southern Africa at 8,000 14C years ago seems to have been generally quite similar to today, although slightly wetter in some areas and slightly drier in others (L. Scott, pers. comm., March 1990). Between 7,000 and 6,000 years ago, micromammal evidence indicates conditions in the Namib Desert were moister than at present (Avery 1991), but these data do not appear to extend back as far as 8,000 years ago.
Madagascar. Moister but cooler than present? Early Holocene evidence from Madagascar is largely lacking. Possible clues come from a pollen diagram that starts in the mid-Holocene, showing slightly moister than present conditions in western Madagascar (Burley 1993). Recently obtained pollen evidence from the Malagasy Plateau in the north-central part of the island suggests a persistence of cooler-than-present conditions (e.g. presence of Ericaceae) during the early Holocene, although the vegetation interpretation is complicated by the change from lake to swamp conditions at this site (D. Williamson pers. comm., Jan 1995).
5,000 14C years ago.
By 5,000 years ago, African climates and vegetation in most areas were drier than in the early Holocene, but still generally moister than today's.
The Levant. Becoming drier. The vegetation map of van Zeist & Bottema (1988) for 4,000 years ago (which we have taken as also likely to be representative of 5,000 years ago) shows that conditions were slightly drier than at 8,000 years ago but with a 70 km band of closed forest extending parallel to the coast, which is perhaps what the present vegetation would be without extensive human interference. This conclusion was based on several pollen localities.
Arabian Peninsula. Becoming drier. Conditions were drier than at 8,000 years ago, but still moister than today. The occurrence of Neolithic sites within Arabia between 5,000 and 3,500 years ago indicates moister than present conditions (Petit-Maire et al. 1994; S. Stokes, pers. comm. Aug. 1994), and Lioubimtseva (1995) suggests that scrub would still have been widespread in the west Arabian mountains and steppe in northern Arabia. On the basis of various indicators, Whitney et al. (1983) suggested that in much of the Arabian Peninsula the annual rainfall was about 250-300 mm during the mid Holocene, in contrast to the present 50-100mm, and enough to give a semi-desert rather than the present desert vegetation.
Saharan region. Drying under way. A switch towards drier climates apparently began around 6,100 14C years ago and intensified around 4,500 14C years ago. There is a fall in the level of many lakes at around 5,000 years ago (Lioubimtseva 1995) reaching a minimum sometime between 5,500 and 4,000 years ago (Damanti & Harrison 1995). A range of other indicators also suggest that by 5,000 years ago, a significant drying of the climate of the whole Saharan region had already occurred, relative to the situation at 8,000 years ago (Lioubimtseva 1995, Vernet 1995). However, the climate was still much moister than at present; there are many indications that rainfall was sufficient to allow a much more extensive vegetation cover than at present.
Baumhauer (1995) suggests on faunal, pedological and lake-level evidence that at his central Saharan site, annual rainfall during the mid-Holocene (after about 6,500 14C years ago) was about 150-250 mm; enough to support semi-desert vegetation. In the present eastern Sahara hyperarid region at Oyo (19N, 16E), Ritchie (1994) suggests on the basis of pollen evidence a cover of steppe and semi-desert communities between 6,000 and 4,500 years ago. This stands in contrast with the present landscape which is almost totally devoid of vegetation. Neumann et al. (1995) have obtained pollen cores from Oursi (14N, 0W) in the western Sahara/sahelian zone. Combining these with the charcoal evidence, they suggest that during the mid-Holocene instead of the present Sahelian vegetation there was a no-present analogue mosaic of denser vegetation communities, some grassy and some more wooded. At that time it seems, the Sahelian vegetation belt apparently did not exist anywhere in its present form, and that it is a product of climatic drying and human impact over the last 3,000 years (Neumann et al. 1995). Despite the no-present-analogue nature of past Holocene vegetation, it is necessary to use some sort of categories for useful mapping, and what are thought to be their nearest modern-day equivalents are presented in my map here, strongly influenced by the positions of the vegetation belts reconstructed by Lézine (1989) for the eastern Sahara on the basis of scattered plant fossil evidence.
Conditions across the Sahara region and the Arabian Peninsula at 6,00014C y.a. (which may in general resemble those at 5,00014C y.a.) have been summarized in a 1-degree database and set of biome maps presented by Hoelzmann et al. (1998), using the extensive pollen and charcoal data from the region. Their map suggests a picture similar to that given in the maps here; note however that from their useage of categories for the present-actual map, their category of 'steppe' appears to correspond to 'semi-desert' in the QEN vegetation scheme, and their 'savanna' corresponds more closely to the QEN 'grasslands' and 'scrub'. Hoezelmann et al. also suggest a very extensive area of wetlands south-east of Lake Mega-Chad, rivalling the lake itself in scale; they suggest that rainfall in the catchment area was around 300-350mm higher than today in order to sustain this high water level. Other extensive wetland areas are suggested for the eastern Arabian Peninsula
Southern and Eastern Africa. Similar to today. Generally, the signs from the many pollen and palaeoenvironmental records across southern and eastern Africa are of mid-Holocene vegetation and climates that were quite similar to those of today, but with a vegetation structure not subject to the intense degradation by domesticated grazing animals that it undergoes at present in some regions. Of course, the situation as regards overall grazing pressure on vegetation may not have changed much in some places where the natural fauna has simply been replaced by a similar density of herded grazing animals. Elsewhere, higher-than-present populations of such large herbivores as elephants may have caused less arboreal vegetation to be present than is now the case (Kortlandt 1984).
Rainforest zone still wetter than now? The indicators cited above for 8,000 years ago also seem to indicate moister, more forested conditions than at present at 5,000 years ago. North of the equator at least, monsoon rains were still much greater than at present (though diminished relative to 8,000 years ago), as shown by the Saharan evidence. From the fact that more Podocarpus pollen than at present is seen in several pollen diagrams before about 5,000 years ago, Hamilton & Taylor (1991) suggest that the climates were still moister than at present.
Slightly more woodland cover in East Africa. Pollen in Lake Victoria around 5,000 years ago seems to indicate a greater-than-present tree cover relative to grass cover, which may be due to a moister climate or perhaps weaker human influence. The general view of slightly moister-than-present conditions with more woodland is corroborated by other workers, such as Maitima (1991) who finds that in the lowlands of the central rift valley there was a drying after 6,500 years ago, with loss of woody vegetation and replacement by grasslands, but only giving the mainly grassland vegetation similar to today's by around 4,000 years ago. In east Africa, lake level evidence generally seems to suggest moister-than-present climates in the Ethiopean rift region and the Ethiopean Plateau, but relatively similar-to-present conditions in the region of the western rift and Lake Victoria (Hamilton 1982) although many lakes do still seem to have slightly higher than today at around 5,000 years ago (Hamilton 1982).
Similar to present in east African highlands. In the east African mountains, pollen diagrams generally indicate vegetation and climates very similar to today's by 5,000 years ago, although Mt. Elgon seems to have had a slightly wetter climate (Hamilton & Taylor 1991). In Kashiru bog in Burundi, at 5,000 years ago there was an increased montane forest cover than at present, in a phase of relatively warm and wet conditions lasting between at least 7,000 and 4,500 years ago (N.B. there is a gap in the data before 7,000 years ago; perhaps due to greater aridity?) (Bonnefille & Riollet 1988).
Southern Africa.
Slightly warmer and moister? In southern Africa, slightly warmer conditions than at present seem to have occurred around 5,000 14C years ago (at the end of a phase lasting from 7,000 to 5,000 years ago), when it was also slightly moister along the Cape Coast (Deacon 1990). However, in general the vegetation cover seems to have been not very different from the present-potential (Burney et al. 1994).
Madagascar. More woodland and forest? Pollen deposits from a lake in south-western Madagascar that date back only during the past 5,000 years indicate a shift from an earlier forest and woodland vegetation (of a type still fairly typical of much of the west side of the Island further north than this particular site) towards somewhat drier conditions with wooded savanna, followed several thousand years later by human influence through burning (Burley 1993).
Agriculture present, but not a major influence on vegetation? At 5,000 14C years ago, agriculture may have been present but was apparently not yet a significant influence on vegetation in Africa south of the Sahara. There have been various reports of possible agricultural vegetation disturbance in the early Holocene of east Africa (back as far as about 8,000 years ago) (D. Olago, May 1994), but the first generally accepted sign of agriculture in subsaharan Africa is a rise in oil palm pollen in the equatorial belt around 3,500-3,000 years ago (Tallis 1990). Significant forest clearance is generally seen as beginning in east Africa at 2,200 years ago in Uganda and 1,800 years ago in Ethiopea. However, some herding and crop growing was taking place in northern Africa and the Levant, though probably with little overall impact on the vegetation. In Madagascar, humans do not seem to have arrived until around 1,000 years ago (Matsumoto & Burney 1994).
Names and current addresses of QEN participating experts (named in the text above) who have made direct contributions to this work on Africa:
R. Bonnefille, Laboratoire de Géologie du Quarternaire, CEREGE, Europole de l'Arbois, B.P. 80, F-13545, Aix-en-Provence Cedex 04, France..
R.M. Cowling, Department of Botany, University of Capetown, Private Bag Rondebosch 7700, South Africa.
H.J. Deacon, Department of Archaeology, University of Stellenbosch, Stellenbosch 7600, South Africa.
H. Elenga, Laboratoire de Géologie du Quarternaire, CEREGE, Europole de l'Arbois, B.P. 80, F-13545, Aix-en-Provence Cedex 04, France..
J.R. Flenley, Department of Geography, Massey University, Palmerston North, New Zealand.
A. Hamilton, World Wide Fund For Nature, Panda House, Godalming, Surrey GU7 1XR, UK.
H. Hooghiemstra, Hugo de Vries Laboratory, University of Amsterdam, Kruislaam 318, 1098 SM, Netherlands.
N. Lancaster, Department of Geology, Arizona State University, Tempe, Arizona 85287-1404, USA.
A-M. Lézine, ORSTOM, Centre de Bondy, 72 Rte d'Aulnay, 93143 Bondy cedex, France.
E.U. Lioubimtseva, Department of Geoecology and World Physical Geography, Moscow State University, Moscow 119899, Russia.
J. Maley, ORSTOM, Laboratoire de Palynologie, Universite des Sciences et Techniques du Languedoc, Montpellier, France
K. Neumann, Botanisches Institut, J.W. Goethe Universitat, Frankfurt am Main, Germany.
D. Olago, School of Geography, University of Oxford, Oxford OX1 3TB, UK.
N. Petit-Maire, Laboratoire de Géologie du Quarternaire, CEREGE, Europole de l'Arbois, B.P. 80, F-13545, Aix-en-Provence Cedex 04, France..
T.C. Partridge, Transvaal Museum, P.O. Box 413, Pretoria 001, South Africa.
F.A. Perrott, Department of Geography, University of Swansea, UK.
N. Roberts, Department of Geography, Loughborough University, UK
L. Scott, Department of Botany, University of the Orange Free State, PO Box 339, Bloemfontein 9300, South Africa.
D. Schwartz, ORSTOM, B.P 1286, Pointe Noire, Republique du Congo.
S. Stokes, School of Geography, 1 Mansfield Road, University of Oxford OX1 3TB, UK
D. Taylor, School of Geography and Earth Processes, University of Hull, Hull HU6 7RX, UK.
M. M. Umer, Department of Geology and Geophysics, Addis Abeba University, Addis Abeba, Ethiopia, P.O.B. 1176.
A.A. Velichko, Institute of Palaeogeography, Russian Academy of Sciences, Moscow.
A. Vincens, Laboratoire de Géologie du Quaternaire, CNRS Luminy, Marseille, France.
E.M. van Zindern Bakker, P.O. Box 1292, Somerset West, 7130, South Africa.
D. Williamson, Laboratoire de Géologie du Quarternaire, CEREGE, Europole de l'Arbois, B.P. 80, F-13545, Aix-en-Provence Cedex 04, France.
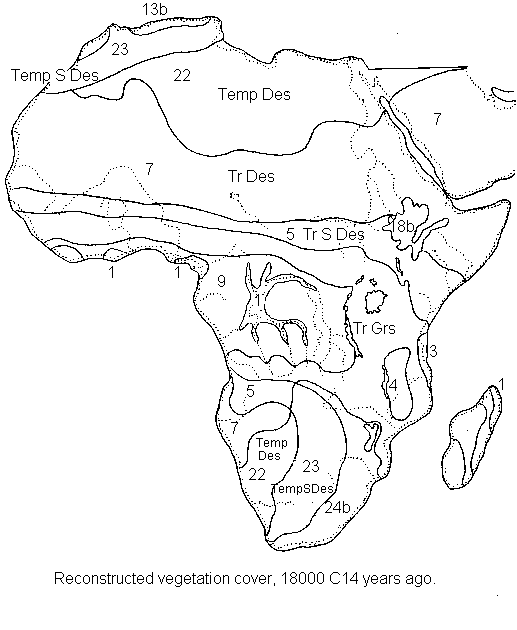 Africa 18,000 14C years ago
Africa 18,000 14C years ago Africa
8,000 14C years ago
Africa
8,000 14C years ago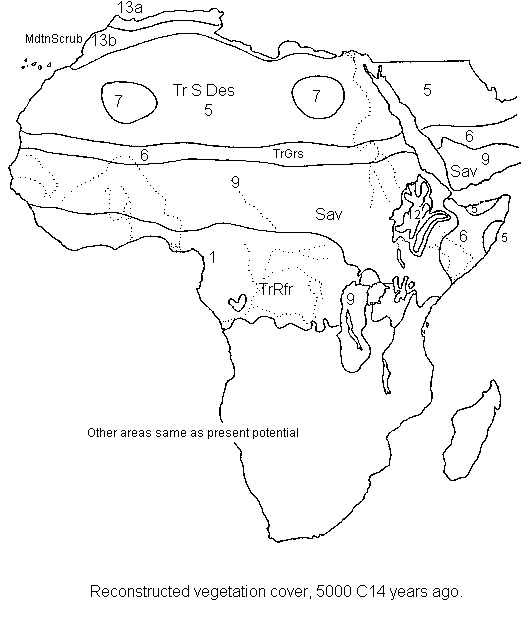 Africa
5,000 14C years ago
Africa
5,000 14C years ago{kind=link}