
 South America
South America
Summary
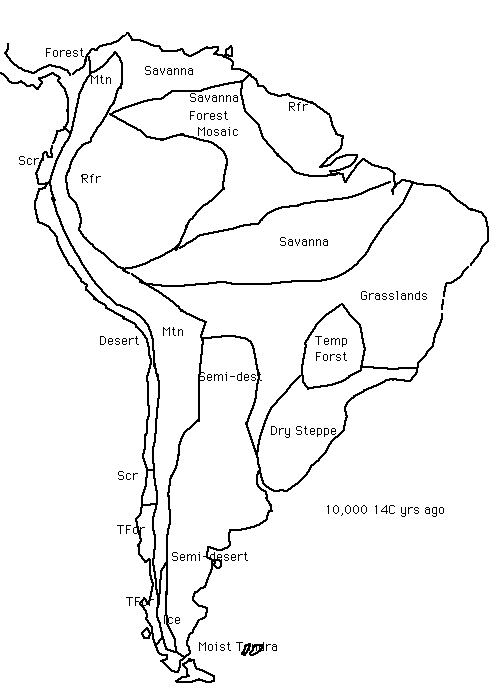
The Quaternary history of South America is generally quite poorly known, although the record is improving rapidly. In general, it would seem that 150-130,000 y.a. the continent showed the general glacial-age pattern of colder and more arid conditions. After about 130,000 y.a., climate warmed and moistened and the forests reached a similar area to the present. After 115,000 y.a., cold and aridity began to influence the vegetation, to an arid, cool maximum around 70,000 y.a., followed by erratic but generally fairly cool and drier-than-present conditions throughout the continent. A second cold, arid maximum began around 22,000 years ago and lasted until about 14,000 14C y.a., after which rainfall and temperatures increased and the forests returned over several thousand years.
Key to the vegetation classification system used in the paleovegetation maps
This is roughly the situation that existed about 500 years ago, when Europeans first arrived. A large area of tropical rainforest occurs in the Amazon and Orinoco Basins, and in scattered areas in the foothills of the Andes and to the south-east where climates are warm and moist enough. Much of the rest of South America has rather dry climates with scrub, grassland or semi-desert. One small area of very moist climates occurs at the southwestern tip, where there are temperate rainforests. Temperate climates also exist at mid-altitudes in the Andes, where there was quite extensive agriculture associated with the Inca civilization
28,000-25,000 14C y.a.; shortly before Last Glacial Maximum. This seems to have been a phase of increasing aridity in Amazonia. Moist conditions in south-eastern Amazonia (the Carajas site) ended about 28,000 14C y.a. Pollen evidence suggests that the forest began to give way to savanna, although it did not finally disappear from the site until about 20,000 14C y.a. (Siffedine et al. 1992).
As mentioned above, a large 'refuge' area of forest seems to have remained in central and western Amazonia, confirmed by pollen evidence published recently by Colinvaux's group (Colinveaux et al. 1996, De Oliviera et al., 1995), from a site in the upper Rio Negro area of central Amazonia. The work by Colinvaux's group is also summarized in this linked web page. At the upper Rio Negro site, forest tree percentages in the pollen record indicate closed tropical forest, with a significant element of lower montane species suggesting cooling. The only evidence for possibly greater aridity in these cores is a slight lowering of water levels of the lakes that were cored. It is important to note that, contrary to some newspaper reports, this latest pollen site does not confirm that the Amazon forest remained intact across the whole region during the last glacial period; what it does show is that a present high-rainfall area that was generally suspected as having remained a forest refugium (e.g. by Clapperton, van der Hammen etc.) was indeed a refugium at the LGM. Broad scale precipitation patterns in the present-day Amazon-Orinoco region show a rainfall minimum extending northwest-to-southeast, and on the reasonable assumption that the distribution of relatively high and low rainfall areas remained much as it is today, the general lowering of average precipitation indicated by several sites would have produced a broad savanna corridor across this region. There would also have a large contraction of the rainforest inwards from the drier northern and southern edges of Amazonia (Clapperton 1993), as is indicated by more direct evidence. The Amazon fan, receiving sediments washed down from the Amazon Basin, shows little palynological or other evidence of increased savanna cover (Haberle 1995), suggesting only minor forest reductions during the Last Glacial period. However it is possible that this record is to some extent biased by gallery forests remaining along the Amazon river course during the last glacial (Kerr 1996).
The maximum aridity in the Amazon region may in fact have been reached around 15,000 14C years ago, based on the evidence from the Huascaran glacier core, which records nitrate levels (an indicator of forest cover) during the last 20,000 years (Thompson et al. 1995, and L. Thompson pers. comm. 1997). Generally, nitrate levels in the glacier are much lower than present throughout the period 22,000-12,000 y.a., suggesting a significantly reduced forest extent across the Amazon region during the last Glacial.
Thus, the signs are contradictory, with the balance of different types of evidence perhaps tending to point to a major forest reduction in from the drier edges of Amazonia, leaving a core area in western and central Amazonia at least. The issue of last glacial forest extent will hopefully be resolved more clearly over the next few years as more sites across the interior of Amazonia are located.
Sedimentological, plant fossil and present zoogeographical patterns (Pregill & Olson 1981) support the view that conditions were markedly more arid in the Caribbean at the LGM, with a reduction in forest and its replacement by scrub and grasslands.
In parts of the present forest-savanna zone of Venezuela (Leyden 1985) desert and semi-desert conditions seem to have existed. Desert areas with shifting sand also seem to have existed in western and north-western Argentina (Iriondo & Garcia 1993, Clapperton 1993), and desert dust was blown eastwards to form extensive loess deposits.
What is presently the moist temperate steppe and subtropical/warm temperate forest zone of eastern Argentina and southern Brazil seems to have been markedly cooler and drier than at present (Iriondo & Garcia 1993). For example, various pollen sites from the Coastal Plain of southernmost Brazil suggest semi-arid conditions with a rather sparse dry steppe after 23,000 14C y.a., ending around 11,000 y.a. (Das Neves & Lorscheitter 1991). For example at a site 20 deg.S, 43 deg.W, the present semi-deciduous forests and woodlands were replaced by grasslands during the last glacial, and warm temperate trees existing only as gallery forests along river sides. Behling & Lichte (in press) find pollen evidence of considerably cooler and drier conditions than present in tropical SE Brazil between about 48,000 and 17,000 14C y.a. (with a slight moistening of climate - indicated by slightly higher proportions of tree pollen - between 17,000 and 10,000 14C y.a.). A cooling of the mean annual temperature by at least 5-7 deg.C across tropical and subtropical SE Brazil is suggested, corresponding to a movement of temperature zones at least 750 km equatorwards, and general replacement of both lowland and highland forests in the Atlantic coastal region by grasslands, throughout the glacial period (Behling 1998, Behling & Lichte, in press).
Further south, in Patagonia, a dry, sparse tundra was present (Markgraf 1993) and the Andean temperate forest was reduced to scattered remnants on the western side of Cordillera (Markgraf 1993).
Helmens et al. (1996) suggest that there may have been a relatively warm, mild period (the 'La Laguna Interstadial') in the high plain of the Colombian Andes at around 19,500-17,500 14C y.a., with partial glacier retreat. The conclusions are based on palynological, palaeopedological and glacial geomorphological evidence. They seem to stand in contradiction to other sources from the area, such as the Huascaran core (see above).
More information and discussion of this time slice is given in the main QEN review.
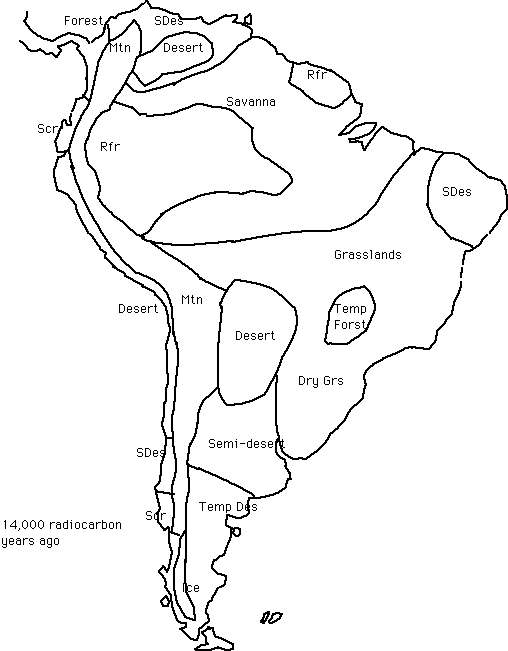
South America 18,000-15,000 14C years ago
Key to the vegetation classification system used in the paleovegetation maps
Conditions the (Pacific) NW coastal plain of South America seem to have been been moister than present, according to a fossil avifauna from about 14,000 14C y.a. (Campbell 1982). Campbell argues that more frequent (annual?) El Ni�o events have caused these moister conditions.
South America before the onset of interglacial climates - 14,000 to 13,000 y.a. 14C ya
Key to the vegetation classification system used in the paleovegetation maps
12,000 radiocarbon years ago. In some parts of South America, somewhat moister conditions become evident at around this time. In Tierra del Fuego, the sparse tundra was replaced by steppe, and the temperate rainforest of the western Andes began to return (Markgraf 1993). At Salitre in seasonal south-central Brazil, forest began to increase in abundance (Ledru 1992) indicating an onset of moister conditions. Forest was present from at least 12,500 14C y.a, on an island at the mouth of the Amazon (Behling 1996), but with certain glacial montane elements (Podocarpus, Ilex) still abundant within the forest.
At the Carajas site in S.E. Amazonia the pollen of rainforest plants began to return, although the forest patches apparently formed an open mosaic with savanna (Siffedine et al., Absy et al.). At Lake Valencia in northern Venezuela, savanna vegetation had replaced the earlier semi-desert, but forest vegetation had not yet appeared (Leyden 1985).
In central America (Costa Rica) the upper forest limits in the mountains seem to have remained at relatively low altitude (2300-2600m) 14,000-11,000 14C y.a. (Islebe & Hooghiemstra 1997). This might have been due to cooler-than-present climates, or some sort of lag in ecological succession.
In the present Araucaria warm temperate forest area in the uplands of of south-eastern Brazil, pollen evidence (24 deg.S, 50 deg. W) beginning around 12,500 14C y.a. suggests a drier, 3-5 deg.C cooler climate, with a vegetation resembling the present moist grassland but with only very scattered areas of Aruacaria forest surviving in the valleys (Behling 1997).
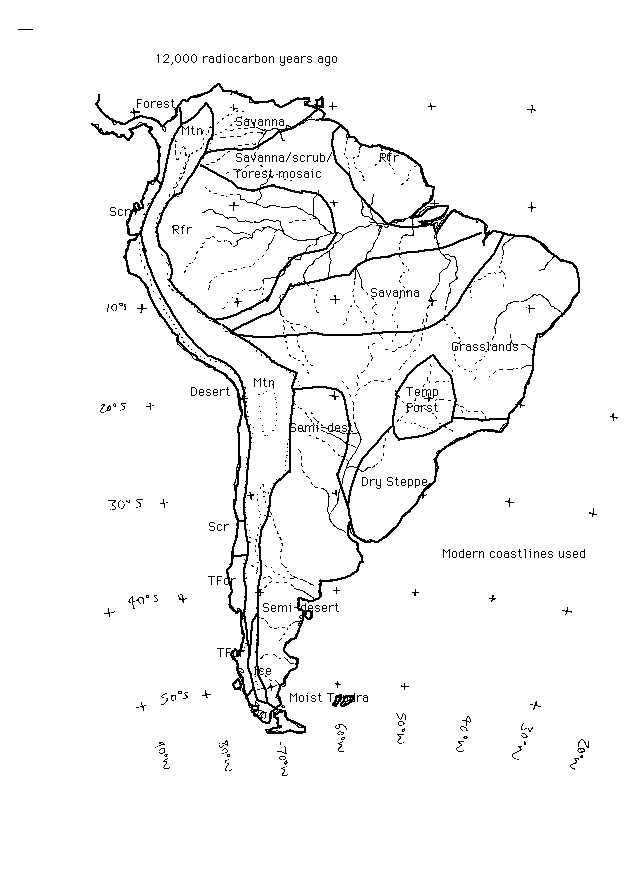
South America during the last glacial - 12,000 14C ya
Key to the vegetation classification system used in the paleovegetation maps
By around 11,500 14C y.a. (when the pollen sequence starts) there was a rainforest cover unusually rich (2% of the pollen) in the montane tree Podocarpus, close the mouth of the Amazon (0 deg.46' S, 47 deg.51'W) (Behling 1997). This forest cover remained similar in composition through the interval of the Younger Dryas, with Podocarpus dying out in the area around 9,500 14C y.a.
It is not presently known whether the forest cover or composition of the main Amazon Basin region was affected by this 'Younger Dryas' climate event, but it is reasonable to suppose that they were influenced in some way.
Maslin & Burns (2000) have studied planktonic oxygen isotopes in the area of Amazon River discharge, and suggest that (from the relative lack of a freshwater oxygen isotope ratio)the Amazon's discharge during the Younger Dryas was at least 40% less than at present. This seems to confirm that the Younger Dryas was a very significant event in the Amazon region, presumably having considerable impact on the returning forest cover (and tending to favour more open vegetation).
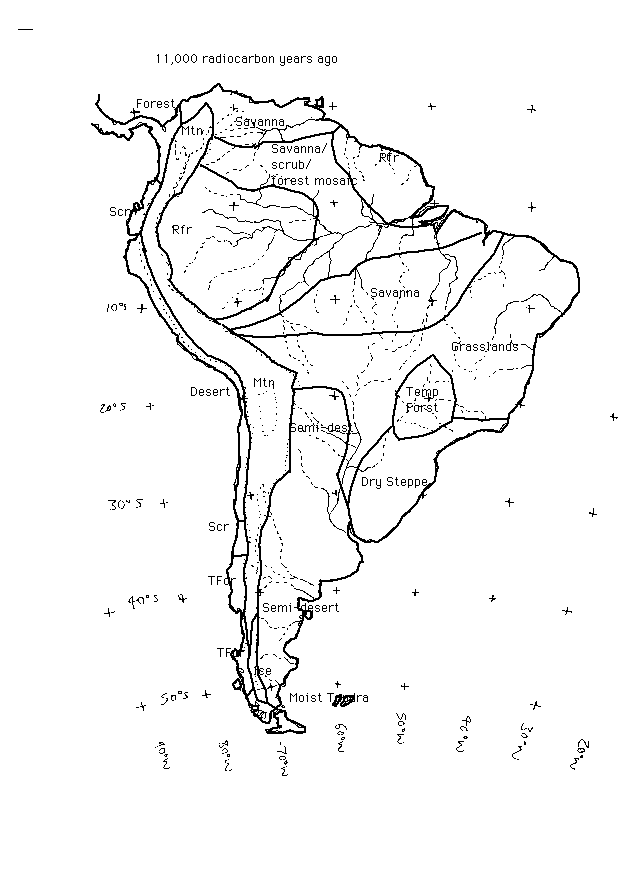
South America during the last glacial - 11,000 14C ya
Key to the vegetation classification system used in the paleovegetation maps
In a pollen core from rainforest near the Amazon River mouth, Behling (1997) finds an initial rise in charcoal and grass pollen, perhaps indicating first colonization and localized forest disturbance in the area by indians.
From detailed study of a highland lake in Costa Rica, Islebe & Hooghiemstra (1997) suggest that the transition from cooler 'Younger Dryas' to Holocene conditions occurred much more gradually than the sudden change observed in the north Atlantic region.
In the south-eastern highlands of Brazil (24 deg.S, 50 deg. W), Araucaria forest gradually expanded during the lower Holocene under cooler, moister conditions to give greater forest cover than at present (Behling in press), and rainforest species expanded in valleys later on. Forest cover in some areas of the semideciduous forest zone (around 20 deg.S) at lower elevations also seems to have increased substantially around this time (Behling & Lichte, in press). However, Behling (1995) suggests that elsewhere in the foothills of the southeastern highlands (17 deg.E, 42 deg.w), in the present semideciduous forest zone, the climate was drier than present throughout the early and mid Holocene, with forest restricted to valleys.In the central and southern Pampa region of Argentina, at around 10,500 14C y.a., several pollen cores (in the area between about 58-62 deg.W and 35-38 deg.S) suggest that there was a substantial shift in the vegetation from a previous dry steppic environment to moist steppe conditions similar to today's (Prieto 1996).
South America during the last glacial - 10,000 14C ya
Key to the vegetation classification system used in the paleovegetation maps
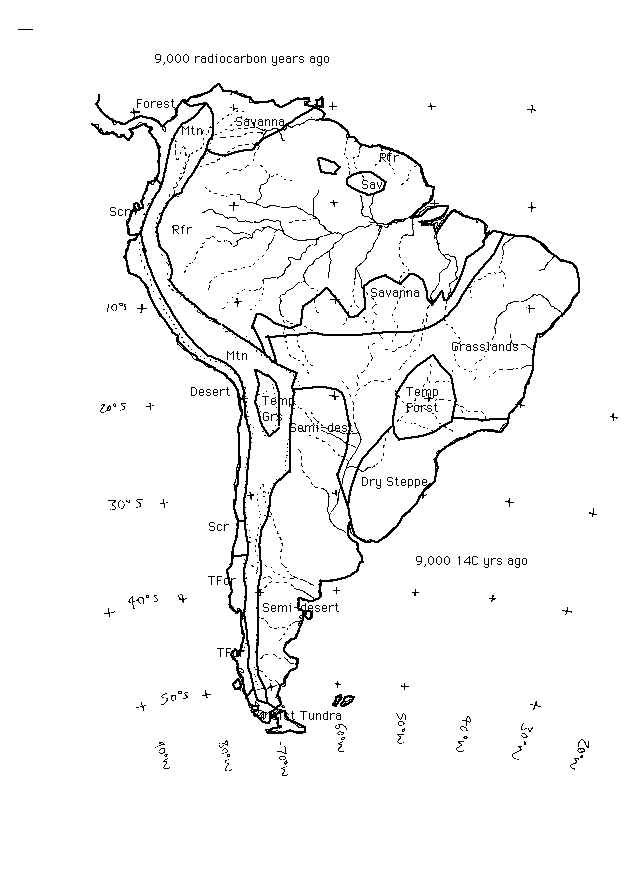
9,000 - 9,500 radiocarbon years ago. At the Lake Valencia site in northern Venezuela, lowland rainforest had reached its maximum coverage (Leyden 1985). In the mountains of central America, forests seem to have reached their present altitudinal limits (3000-3300m) between around 9,000 and 8,500 14C y.a., and forest cover in the lowlands of Panama and Guatamala may also have been incomplete until around this time (Islebe & Hooghiemstra 1997). Likewise, at sites in the present rainforest zone of Amazonia, closed forest seems to have reached full Holocene extent by around 9,000 radiocarbon years ago. However, nitrate levels (an indicator of forest cover) in the Huascaran Glacier in the equatorial Andes, suggest that forest recovery in the Amazon region was delayed and did not reach its full Holocene extent until around 8,000 years ago (Thompson et al. 1995).
Grass pollen (suggesting open ground) and small charcoal particles in sediments begin to appear in abundance at a site in the rainforest near the mouth of the Amazon at around 9,500 14C y.a., and have been taken to suggest the onset of significant disturbance by indians along the shores of the lake where the core is taken (Behling 1997).
Further to the south, in subtropical Brazil and in Argentina, there are contradictory signs of the timing of ecological and climatic changes. For example at Salitre in south-central Brazil, conditions seem to have been somewhat cooler than at present, though with forest having returned (Ledru 1992). Drier-than-present conditions - with less forest than during the late Holocene - may have existed in the subtropical forest area of SE Brazil from about 8,000 years ago until about 3,000 years ago, indicated by charcoal phases followed by savanna incursion (Vernet et al. 1994). Other evidence of more arid than present conditions at the 8,000 y.a. timeslice comes from a pollen record at Lago do Pires; 17 deg.57'S, 42 deg. 13'W (Behling 1995b). A herb savanna seems to have predominated, with gallery forests in the valleys, in contrast with the latest Holocene when closed semideciduous forest covered the whole area. A similar pattern is found by Das Neves & Lorscheitter (1991), with forest appearing about 4,000 14C y.a. at a coastal plain site at 29 deg.S, 50 deg.W in SE Brazil.
In Argentina, the climate may also have remained in a relatively cold, arid mode than its later Holocene state (Iriondo & Garcia 1993), with desert-like conditions in the Chaco region. The final climate switch to 'optimum' (moistest and warmest) conditions may have occurred at around 8,000 radiocarbon years ago, and to have lasted until about 5,000 14C y.a. after which there was a return to rather arid conditions. However, according to Prieto (1996), the initial switch to moister-than-present conditions in the Pampa and elsewhere began considerably earlier, at the start of the Holocene. Prieto suggests, on the basis of work by Dillehay et al., that Iriondo & Garcia were relying on unreliable radiocarbon dates. He also notes work by Tonni et al. which suggests a switchover in the vertebrate fauna away from Patagonian and central Argentinian species towards Brazilian species of mammals at the start of the Holocene.
In the extreme south, the Valdivian temperate evergreen forests had returned on the western side of the Andes but had not yet spread through to the eastern side (Markgraf 1993).
South America during the early Holocene - 9,000 14C ya
Key to the vegetation classification system used in the paleovegetation maps
After 7,000 14C years ago. A relatively dry phase seems to have occurred in Panama, Guatamala and Costa Rica between about 7,000 and 5,000 14C y.a., with a slight opening up of forest to savanna (Islebe & Hooghiemstra 1997).
There is evidence from both the Colombian Andes (lowering of altitudinal forest limits) and sub-tropical Chile, of cooler, moister conditions around 2,600 14C y.a. (2,600 y.a.). A dry phase also seems to have occurred on Haiti between about 2,600 and 1,500 y.a. (evidence summarized by van Geel et al. 1996).
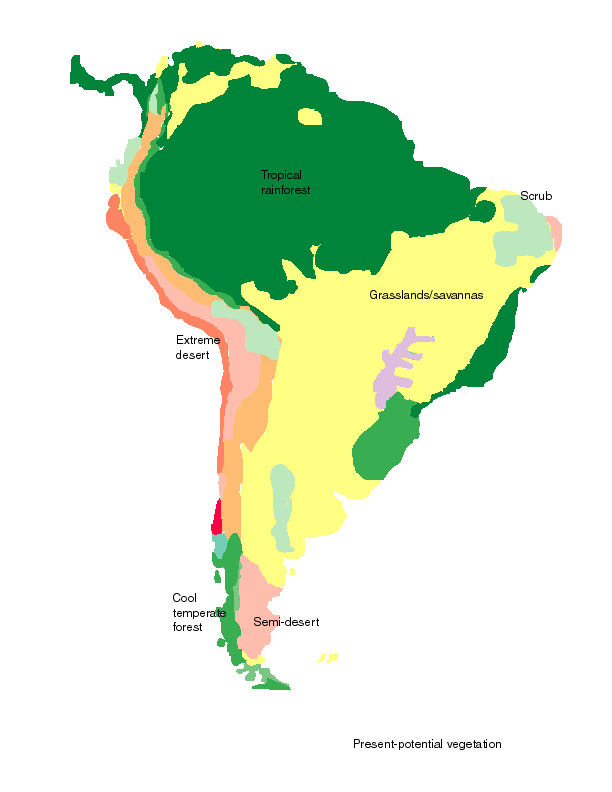 Present potential vegetation
Present potential vegetation{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}