
Last modified Sunday 13th April 1997
SOUTH & CENTRAL AMERICA including Mexico and the Caribbean
Return to QEN Home Page RETURN TO ATLAS MAIN MENU
For Central America and Caribbean region, see:
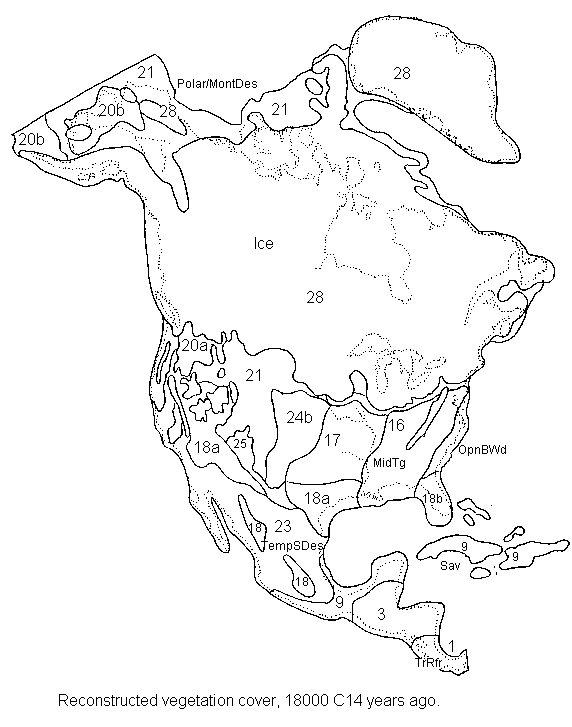 North
& Central America 18,000 14C years ago
North
& Central America 18,000 14C years ago
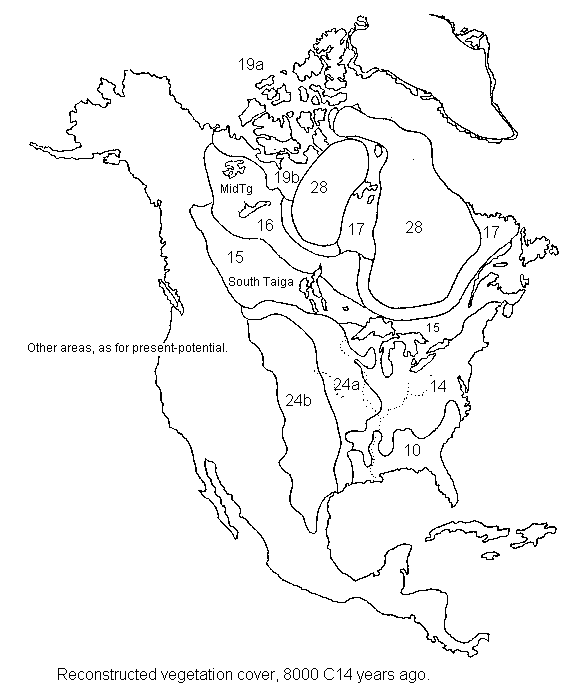 North
& Central America 8,000 14C years ago
North
& Central America 8,000 14C years ago
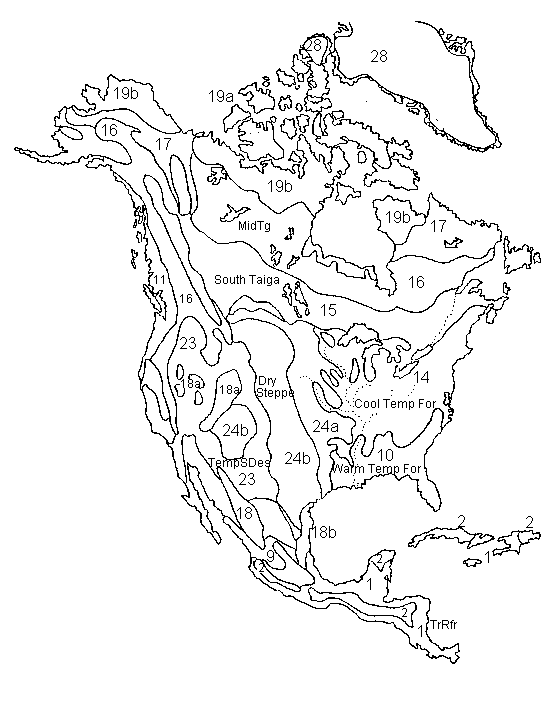 North
& Central America 5,000 14C years ago and present-potential
North
& Central America 5,000 14C years ago and present-potential
For South America, see:
 South
America 18,000 14C years ago
South
America 18,000 14C years ago
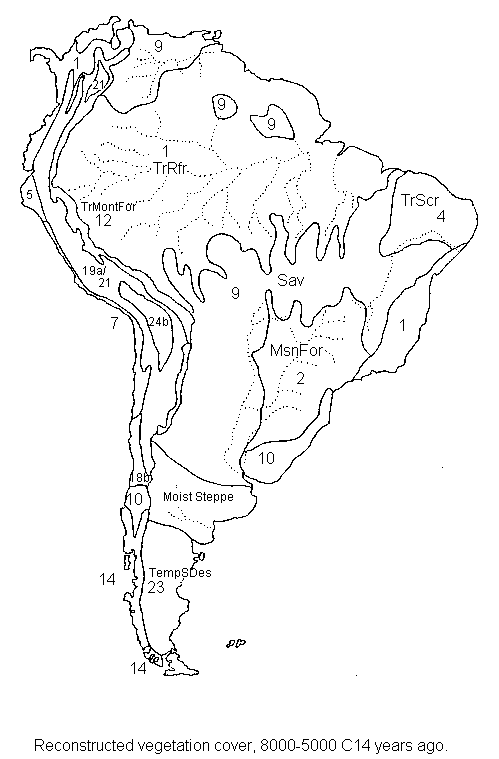 South
America 8,000 and 5,000 14C years ago, combined
South
America 8,000 and 5,000 14C years ago, combined
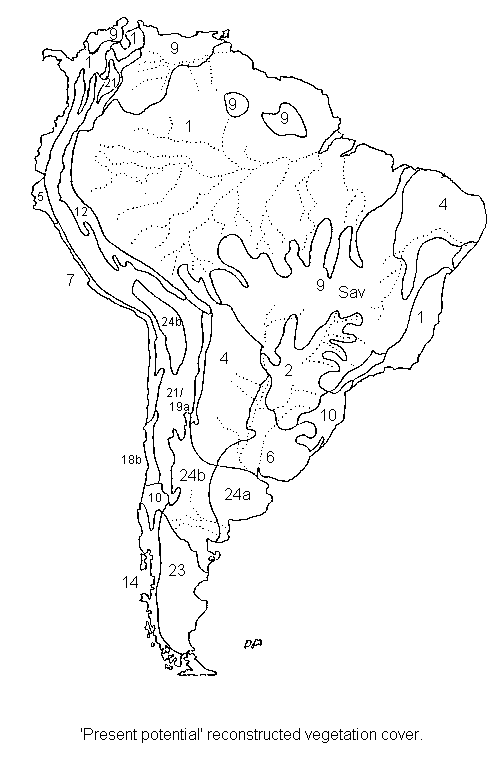 South
America, present-potential.
South
America, present-potential.
VIEW NEW SET OF PALAEOVEGETATION MAPS (PRELIMINARY) AT MORE FREQUENT TIME INTERVALS
List of References (separate document)
Key to the vegetation classification system used in the atlas
18,000 14C years ago (Last Glacial Maximum, or LGM). .
Note that a number of new sites (referred to below) have come to our attention since this diagram was made, and these sites are not shown on this map.
An ice sheet was present at the southern end of the Andes, flowing down onto the exposed continental shelf. Other ice caps were present in the high Andes further north. Ice sheet extent in this map is based on map drawings by Denton & Hughes (1991), Iriondo (1988), and L. & C. Heusser (pers. comm., Feb. 1990). In the Bolivian Andes, the maximum glacier advances seem to have been around 20,000-18,000 radiocarbon years ago, coinciding with the 'global' glacial maximum. However, in the Ecuadorian Andes the maximum was around 25,000 years ago, whilst in the Mexican uplands the maximum glacier extent occurred about 15,000 years ago (Heine 1995).
Broad-scale climatological evidence. Recent broad-scale evidence from ice cores in the high Andean Huascaran Glacier core, (10 S, 78 W) indicates a much greater dust input (about 200 times Holocene levels) between 20,000 and 14,500 years ago (Thompson et al. 1995). This favours the view that at least some areas in the South American tropical region were considerably drier than at present during the LGM. Broecker (1995) suggests that the dust input reflects severe Amazon lowland aridity, although one should probably consider instead that the dust was derived from the desert areas that had appeared outside the main Amazon Basin (see below).
Nitrate levels in the Huascaren Glacier fell by a factor of two to three and dust concentrations increased 200-fold. Thompson et al. conclude that these indicate that there was substantial drying and a significant reduction (40-50%) of the rain forest in the nearby Amazon River basin, with forest cover being much less than present at the LGM, but reaching a minimum slightly later (about 15,000 14C y.a,).
Hydrogen isotopes in the same glacier cores suggest that sea surface temperatures in the tropical Atlantic source area for the moisture were some 5 or 6°C cooler than at present (contrasting with previous, planktonically-based estimates which suggest at 2°C lowering), and oxygen isotopes suggest the same picture for the general region where the ice was being deposited (Thompson et al. 1995, Broecker 1995).
A lake over Amazonia? Generally thought very unlikely. On the basis of the topography and drainage patterns of the Amazon, Campbell (1989) has suggested that a huge lake ('Lake Amazonas') existed in the Amazon Basin at this time, flooding the basin to 300m above sea level. This would have been held in place by a huge sediment dam in the lower Amazon (or possibly through downwarping of the crust in the central Amazon), and its discharge events in the early Holocene may have corresponded to freshwater discharge events recorded for the Amazon delta region. Of course, if Campbell's hypothesis were correct, it would make discussion of the LGM vegetation of the Amazon Basin largely irrelevant because everything would have been covered in water. This view is strongly disputed by most who work on this area. Colinvaux (1993) cites recent work by Rasanen et al. as disproving the supposed existence of Lake Amazonas, as there is no evidence of the widespread layer of sediment that such a lake would have been be expected to produce. Clapperton (1993) discusses the hypothesis at length, before rejecting it completely on various grounds. Note that LGM pollen cores dating from the LGM have also now been obtained from the central Amazon (see below), showing directly that at least some areas were not inundated (though this site is on a hilltop).
Central America and the Caribbean.
Fairly similar conditions to the present in central Mexican uplands? In the highlands of central Mexico, three pollen records indicate a predominance of Pinus-Juniperus woodland at 18,000 14C years ago, fairly similar to the vegetation today (Markgraf 1993). However, the absence of deciduous broad-leaved taxa suggest that the climate was in fact slightly drier than today's. The higher percentages of steppic species such as Artemisia and Ambrosia likewise suggest drier, more open vegetation. From another core from Chalco Lake in Mexico City (part of the highland massif) (19. 30' N, 99. 00' W), aridity around 18,000 y.a. is indicated by the absence of the Holocene and late-glacial forest cover, and the dominance of grassland vegetation (Lozano-Garcia et al. 1993).
At higher altitudes, a combination of geomorphological and palynological evidence suggests a temperature depression of 4.2°C above 4,000m (Heine 1995).
There are not yet any records from the Mexican lowlands dating back to the LGM, but one upland core in coastal mid Mexico suggests a drier vegetation of grassland and herbs just before the start of the Holocene, being replaced by the Holocene vegetation of savanna-forest mosaic (Markgraf 1993).
Colder and drier, with grassland and sparse scrub in lowland Guatemala. In Central America, the evidence is of generally drier than present conditions. Binford et al. (1987) present pollen evidence that indicates the prevalence of savanna before 10,000 14C years ago in the Peten Lowlands of Guatemala, an area which currently supports seasonal semi-evergreen forest. In a lowland site (Lake Quexil) in the same area of Guatemala, Leyden et al. (1993) have found pollen, sedimentological and isotopic data indicating a much drier and much cooler climate than today during the period that includes the LGM. The vegetation was a sparse dry temperate scrub and grassland, of a type that now grows above 1500m. A mean annual temperature depression of about 6.5 - 8.0°C is suggested from the pollen evidence.
Deciduous forest in lowland Panama. In the lowlands of Panama, pollen evidence indicates that forest was still present in the area that presently supports rainforest, but that it was more seasonal (drought deciduous) and more open than today (Bush & Colinvaux 1990). Again, the occurrence of montane tree species suggests significantly cooler temperatures on land, even at sea level, although Bush suggests that maximum cooling may have occurred somewhat after the LGM. The mixture of lower montane and lowland tropical elements suggests a forest community that has no compositional analogue in the present-day world. Piperno et al. (unpublished manuscript) discuss their core from another site at La Yegunda, a lake at 650m a.s.l. (8 N, 8 W). The part of their sequence from the last glacial, beginning at 14,000 years ago, shows a vegetation subject to a substantial reduction in rainfall (about 1000mm), which they suggest would have been enough to reduce the vegetation on the adjacent central plain of Panama (presently natural forest cover) to thorn scrub and savanna parkland. The site itself remained forested however, with the presence of Quercus suggesting a 5°C lower-than-present temperature. Colinvaux (pers. comm., Aug. 1993) appears to suggest that the drying observed in lowland Panama was in fact mainly a lower Holocene event, although he does not specify his reasons for thinking that this is so.
Lowering of treeline in Central American mountains. From pollen evidence, Markgraf (1993) reports that there was a treeline lowering of 600-800m in the Central American mountains, relative to the Holocene. From this, mean temperatures would seem to have been some 4°C below the present. Below 2400m, arboreal taxa were predominant.
However, a more drastic temperature and vegetation zone lowering is suggested by results from high altitudes in Costa Rica, at La Chonta bog in the Cordillera de Talamanca, the upper forest line seems to have been at around 2000m elevation during the LGM, compared to the present forest line of around 3300-3500m (Hooghiemstra et al. 1992). This apparently corresponds to a temperature lowering relative to the present of around 8°C at these altitudes.
Drier conditions in Caribbean. For the Caribbean islands, the general indications from pollen, sediment fluxes, lake levels and biogeography are of drier conditions. Bonnatti & Gartner (1973) obtained a core from the central Caribbean Sea, and noted fluctuations between drier and wetter phases (the dry phases indicated by higher quartz content and lower kaolin content). They suggested from the change in sedimentation at the topmost part of the core, that the dry phases represent glacial periods such as the last glacial. On Haiti (Hodell 1990), the record does not go back to the LGM although during the possibly 'glacial-like' phase of Younger Dryas age, lake levels were lower than today, and in an area naturally occupied by mesic forest there was instead low scrub (consisting of palms and montane shrubs) and grassland - a possible indicator of what LGM conditions were like? Similarly, the changing level of a karstic lake in western Jamaica would seem to indicate drier conditions than at present during the late glacial than during the Holocene (poster in School of Geography, Oxford, 1994; J.A. Holmes, F.A. Street-Perrott & A. Perrott; published as Street-Perrott et al. 1993 in J. Palaeoliminology).
Galapagos aridity. There is also lake level evidence for drier LGM conditions on the Galapagos islands, to the west of South America, before the beginning of the Holocene (Colinvaux 1972). Colinvaux (1987) also cites pollen evidence from the Interandean Plateau of Ecuador to suggest that this aridity likewise affected the Pacific coast of South America adjacent to the Galapagos (see below).
The northern coast of South America. Savanna and dunes in Venezuela. To the north of the Amazonian region, close to the northern shore of Venezuela, a pollen record starting at around 13,000 BP shows sparse grassland vegetation in an area which was occupied by open deciduous forests up until pre-agricultural times (Leyden 1985). In the regions of northern Venezuela which are now naturally savanna, pollen evidence going back to about 15,000 years ago indicates considerably more arid than present conditions (Van der Hammen 1984). On the basis of the extensive 'fossilised' sand dunes consisting of wind-worked grains in the llanos region of Venezuela, Clapperton (1993) reconstructs extensive dune fields and sand desert north-west of the Orinoco River. M. Sarnthein (pers. comm., June 1993) has told me that he has recently seen data which confirm his reconstruction of dune fields in southern Venezuela at approximately the LGM, but he does not specify the source of this information.
Drier in Guyana region. For the coastal Guyanas also, Van der Hammen (e.g. Wijmstra and van der Hammen 1966) finds evidence of drier conditions prior to 13,000 14C years ago. During glacial-time low sea-level periods, northern Guyana (Georgetown) and Surinam - presently wet forest - were covered with dry grass savanna.
Reduction in forest cover of Venezuelan highlands. For the Venezuelan Highlands (a lower, northern extension of the Andes), Salgado-Labouriau et al. (1977) have found pollen evidence of a low tree line and cold, dry conditions prior to 12,600 14C years ago. In this site, at around 13,000 years ago when deposition resumes after a hiatus during the last glacial, the level of the modern lake Valencia was reduced to swamp and surrounded by thorn scrub or savanna rather than the present seasonal forest (Thomas & Thorp 1992).
Amazonia.
The region in general. Little is known of large areas of Amazonia at the LGM. The 2-3 fold reduction nitrate input to the Huascaren Andean glacier (10S) (see above) is suggested as indicating that there was much less forest cover in South America during the Last Glacial period, reaching a minimum at around 15,000 radiocarbon years ago (Thompson et al. 1995, Broecker 1995), but the true controls and source areas of this nitrate flux may be open to question.
The peripheral regions and the edges of the Amazon Basin itself often show fossil or sedimentological evidence of drier-than-present climates, but not always. Most of those who work on the Quaternary of the South American tropics believe that the general climatic aridity seen elsewhere in South America would have extended across and drastically reduced the extent of rainforest in the Amazon Basin at the LGM. However, Colinvaux believes that the climate systems of (for example) the western Andes and the Caribbean are sufficiently distinct from that of the Amazon Basin that there may have been little drying of climate and reduction in lowland forest cover. Instead, Colinvaux emphasises cooling as a more important influence on the glacial-age ecology of Amazonia.
Haffer's refugial hypothesis. On the basis of evidence of drier conditions at about the LGM around the periphery of Amazonia, and from available knowledge on diversity patterns of rainforest plants and animals, Haffer (1969) and others suggested that glacial-age aridity reduced the forest to small and scattered refugia, which presently have higher species diversity. However, this hypothesis has been to some extent put in doubt by evidence that the apparent diversity patterns might be a collecting artefact (Nelson et al. 1990) and also evidence that a montane-type forest had replaced the species-rich lowland rainforest in one of Haffer's proposed refugial areas, in the eastern foothills of the Andes (Liu & Colinvaux 1985) (see below).
Pollen evidence of the history of rainforest distribution;
Amazon Delta evidence of forest survival.
A recent source of evidence suggests that although there was considerable cooling at the LGM, Amazon aridity was less severe than some might suggest. A long core (ODP Leg 155) obtained offshore from the Amazon Fan shows an LGM pollen flora with greater-than-present representation of grassland/savanna species, but also with abundant pollen from trees of cool and moist environments, such as Alnus and Podocarpus. This suggests that there was some drying but also a considerable persistence of forest in the Amazon lowlands at the LGM, under cooler (perhaps 6°C lower) conditions than at present (Haberle 1995). However, Kerr (1996) notes that greater aridity and savanna expansion during the last glacial might have been disguised by a high influx of tree pollen into the rivers from gallery forests.
LGM forest survival in central Amazonia. At least one LGM forest refuge has now been found by Colinvaux's group. In four recent lake cores from a hill in the upper Rio Negro area (ON, 67W. 300m a.s.l.) of west-central Amazonia, De Oliveira et al. (1995) and Colinvaux et al. (1996) find pollen evidence for persistence of closed forest at the LGM with a considerable montane element (about 10% Podocarpus). This result seems to confirm that there would have been a considerable area of closed forest in Amazonia, covering at least the current area of higher rainfall in the centre and west of the Amazon Basin. However, its community composition was apparently much altered from today's, indicating a 5-6°C cooling. As we have earlier pointed out on this web page, and as Kerr (1996) also points out, this site is within the current high-rainfall core area of west-central Amazonia (more than 2500mm annual precipitation) and so it would perhaps be expected to have remained as forest even if there had been considerable forest regression in the east, north and south of the Amazon region.
Eastern Amazonia; forest regression from the south-east. It is generally thought that at the LGM the forest would have regressed from areas that are presently the most marginal, due to reduction in rainfall along approximately the same gradients that occur today.
This view seems to gain support from the evidence of loss of forest from the edges of the Amazon Basin and its replacement by grasslands or savannas. In Serra dos Carajas (50W, 6S) in south-eastern Amazonia - a plateau area which presently receives relatively low rainfall, with semi-evergreen rainforest and some open woodland and savanna and annual precipitation around 1,500-2,000 mm (compared to adjacent regions of 2,000-3,000 mm) - lake pollen evidence indicates a savanna expansion, increased clastic content of sediment, and then a sedimentary hiatus at around the LGM, possibly due to aridity causing the lake itself to dry up (B. Turcq, pers. comm.; Absy et al. 1991, and Ledru 1992).
Two more cores taken from swamps in the Carajas area (Sifeddine et al. 1994, Manuscript submitted) agree with this picture in showing a general forest regression and replacement by scrub and savanna, and a lowering of the water level followed by a hiatus at the driest phase constrained by radiocarbon dates to between around 20,000 and 13,000 14C years ago. Note that Colinvaux feels that the vegetation changes indicated in the Carajas area may be unrepresentative of the situation across the Amazon region in general, because it is at the physiological margins of rainforest vegetation. He suggests (pers. comm. 1993) that the main bulk of the Amazon rainforest would have suffered relatively little from such a change in water balance across the region, and not enough to cause much of the forest to give way to woodland or savanna.
Behling (1996) presents results from a lake core from an island in the Amazon delta (0 46' S, 47 51' W) starting at around 11,500 14C y.a. He finds that a diverse closed forest vegetation (with no savanna elements) already predominated at this time, though with abundant montane elements (Podocarpus, Ilex) suggesting a cooler climate. To what extent this vegetation represents the type that was present in the general area several thousand years before at the LGM is uncertain.
N.B. Even the present-day rainfall distribution over the Amazon Basin is still a matter of some conjecture because of the lack of meteorological stations within the forest zone, the considerable year-to-year variability in rainfall, and the strong influence slight differences in local topography on storm precipitation. Different published annual precipitation maps suggest differing distributions of rainfall gradients and maxima, based on different methods of extrapolation (often heavily biased by particular abnormal point values). On the advice of J. Roberts (Institute of Hydrology, Oxfordshire, England; pers. comm. 1991) and P. Jones (University of East Anglia, pers. comm. 1990), the Times World Atlas precipitation map can be seen as generally reliable on a broad scale. This precipitation map is used here in a general sense as a reference point for discussion of rainfall gradients in the past and present.
Southern edge of Amazonia; strong forest regression. At Katira Creek (approx. 8 S, 56 W) in southern Amazonia, in Rondonia, there seems to have been a phase of very open savanna vegetation (95% grass pollen, <5% arboreal pollen) on a grey clay soil that has been radiocarbon dated to around 18,500 yr. ago. (van der Hammen & Absy 1994). This pollen-bearing layer is overlain by 2-6m of a tough red clay that closely resembles the present colluvial deposits of semi-arid seasonally dry parts of the Brazilian Highlands. Thus, the vegetation at Katira at the time the red clay was formed may have been a sparse, treeless semi-arid grassland, not the moister savanna generally assumed for the LGM period, which van der Hammen and Absy suggest was present at around 18,000 radiocarbon years ago (i.e. the LGM by the standard definition). This in turn is followed by another grey clay layer bearing predominantly grass pollen, indicating a return to somewhat moister savanna conditions. On the basis of the evidence of a savanna phase, van der Hammen (pers. comm. June 1992) suggests that during LGM conditions there was about a 1000m lowering of rainfall, converting areas which at present have rainfall below 2000 mm into savanna and savanna woodland, but leaving a substantial core area of forest in central and western Amazonia. However, considering the red clay layer, perhaps this interpretation underestimates the full severity of the arid conditions that occurred during (or perhaps just after) the LGM, for the red-clay layer implies an even greater decrease in rainfall at around this time.
The radiocarbon date at Katira appears to eliminate the argument of Colinvaux et al. (1989) that the lack of precise dating control could just as well allow the savanna phase to have been a Holocene event. They had suggested that the site is within 100 km of present-day patches of savanna, occurring on sandstone massifs, and has a relatively strongly seasonal climate for rainforest. Thus, in their view small Holocene climate fluctuations could easily have allowed savanna to expand over this site. Clapperton (1993) argues against a Holocene age on the grounds that that it is most unlikely that such a thickness of sediment could have accumulated during the Holocene under a predominantly closed vegetation cover. It seems that both on stratigraphic grounds and on the basis of the single radiocarbon date, a very strong LGM-late glacial regression of rainforest in southern Amazonia is incontrovertible.
North-western Amazonia. Van der Hammen et al. (1992) describe a section from the middle Caqueta River in the Colombian Amazon lowlands, in the North-western Amazon (an area that presently has an everwet rainforest climate). Pollen does not occur in the section that includes the LGM (dated 26-13,000 14C years ago), but is present in part of the section roughly dated to 70-40,000 years ago. This earlier section shows savanna pollen and the pollen of trees indicating drier vegetation types, suggesting an increase in semi-open savanna-like vegetation during this earlier (lower Pleniglacial) glacial time. By analogy with the lower Pleniglacial (previous glacial maximum) and upper Pleniglacial (LGM) of the well-dated Carajas core this might be taken to indicate that the pattern of savanna expansion during the upper Pleniglacial (LGM) also extended to this area, albeit with only a weak opening up of the forest. The Carajas core shows a stronger savanna episode during the LGM interval than during the lower Pleniglacial (around 70,000 years ago), and perhaps indicates that the general pattern of dryness in the east would having extended across to western Amazonia as well. However, this is a case of us 'clutching at straws', for lack of any more conclusive evidence to discuss!
Zoological evidence of Amazonian last-glacial conditions. If the LGM rainforest area was in fact much less extensive than at present, one would expect that the (now extinct) savanna grazing fauna of South America would have been present in areas that are now occupied by forest. Indeed, fossils of many species of savanna-adapted grazing mammals (e.g. Eremotherium, Toxodon, Palaeolama, Vicunga, Glyptodon, Pampatherium, Mixotoxodon and Haplomastodon) have been found at exposures scattered along the rivers of western Amazonia. These include sites in the upper Napo River catchment (approx. 1 S, 76 E), presently a hypermoist area and postulated rainforest refugium. A larger number of sites have been found in SW Amazonia (scattered between 8-12 S and 65-75 W) in the Jurua and Ucayali catchments, and well within the present rainforest zone (Latrubesse & Ramonell 1994, Rancy 1991). Other animal fossils from the same sites are from species of riverine or closed forest, which might suggest localised forest patches along rivers in a mainly or partly open grassland environment. Rancy (1991) suggests that at certain stages (probably late Quaternary glacial maxima, according to him) the rainforest all across western Amazonia was fragmented to leave savanna-forest mosaics or savanna with gallery forest. Dating precision of the fossils is often poor and circumstantial, with the position and form of the sediments suggesting a late Quaternary age (Rancy 1991). Several southwestern Amazonia sites have been radiocarbon dated to the period before about 37,000 years ago (Latrubesse & Rancy 1995), suggesting that there had been strong rainforest regression before the LGM (a middle Pleniglacial date is suggested by Latrubesse & Rancy); whether this forest regression recurred or continued up until the LGM seems uncertain from their results. The fact that most South American grazing mammals became extinct after the earliest Holocene certainly suggests that a Last Glacial age for the aridity is more likely than a Holocene age. There is clearly a need for further work on the age of these animal fossils, for they could be crucial to an overall understanding of the stability of the Amazon rainforest during the Quaternary.
Geomorphological evidence of glacial-age climates from Amazonia. There are various signs from geomorphology that conditions in the Amazon Basin were very much drier during the LGM. Some of this evidence is described below. Frustratingly however, there are no precise radiocarbon dates to either prove or disprove these ideas, and as with some of the fossil evidence the basis of these arguments must rest on the general context in which the features are found.
Dune fields in the central Amazon; indicating past phases of seasonal aridity? At some stage during the late Quaternary (not necessarily the LGM), stabilized dunefields formed along the course of the central and lower Amazon under seasonal river level conditions (formed as river floodplain dunes and sand bars) resembling those of present Orinoco River in a savanna climate (Iriondo & Latrubesse 1994). Iriondo & Latrubesse (1994) suggest that at the Last Glacial Maximum there was a long, broad corridor of seasonally exposed dunes in lower Amazon from about 60 deg.W to the river's mouth. In their scenario, wind would have carried the sand and dust from seasonal river bars and sedimentation into surrounding savanna, ultimately giving rise to dune fields.
River downcutting in north-west Amazonia; indicator of aridity? Van der Hammen et al. (1992) describe a section from the Caqueta River in north-western Amazonia (mentioned above). They find that there was an episode of strong downcutting at some stage between 26,000 and 13,000 years ago. They suggest that this could have been caused by aridity (but could lower sea level also be a cause?). A lower river terrace was formed, with sediments lacking in organic matter - possibly an indicator of relatively dry conditions, but this in itself preventing radiocarbon dating.
Gravel deposits in western Amazonia suggesting greatly reduced riverine fluxes. Dumont et al. (1992) studied the deposits of three western Amazonian rivers to make inferences about the paleoclimate of western Amazonia. Gravel-size sediment, up to ten times coarser that the present sandy deposits, is interpreted as indicating irregular fluvial regimes during the Pleistocene glaciations. Paleofluvial morphology from the oldest parts of the Ucayali floodplain is indicative of a discharge (at about 13000 years BP) 15% to 10% of that of the present river. A period of low precipitation which occurred between 4600 and 1000 years BP has also been documented from paleosols and sedimentary deposits. The high stage of the Tapiche river was lower than the present low stage.
Pebbles in western Amazonia. Latrubesse & Ramonell (1994) report possible evidence of Amazon aridity; large pebbles (up to 10 cm in diameter) were deposited by lowland rivers in the SW Amazon Basin. Clapperton (1994) suggests that these indicate a combination of strong precipitation events in the Andes and climatic aridity in lowlands, reaching a maximum at LGM.
Braided-stream sediments in south-western Amazonia; deposited under more seasonal conditions? Clapperton (1993) summarises information and opinion on a section at Beni, in the north of Bolivian Amazonia. He suggests that coarse braided-stream sediments at the base of the section indicate a much more seasonal climate than at present (only fine-grade alluvium is accumulating at present, under much more gentle conditions). Clapperton hypothesises that this seasonal rainfall regime occurred before the beginning of the Holocene, during the glacial interval.
Clapperton (1993) has summarised data on geomorphological features within the present rainforest area (some of which is referred to above) that might be taken to indicate drier conditions and recession of rainforest during the period which included the LGM. Although there is no close dating control (a lack of LGM radiocarbon dates, for instance), Clapperton argues on the basis of the stratigraphic context of these features, and their possible correlation with indications of aridity from other places within and outside Amazonia, that they are evidence of a dramatic reduction in rainforest at the LGM. He suggests that the widespread downcutting of the river under drier conditions of lower sea level, would have produced an 'Amazon canyon' of incised rivers. The resulting lowering of the water table on the lands between the river courses would have further reduced rainforest cover by adversely affecting its water balance, as would the lack of internal 'recycling' of water by evapotranspiration. Thus, in a preliminary sketch map, Clapperton (1993) suggests on the basis of present-day rainfall patterns that there was a general replacement of forest by savanna or scrub at the LGM, with the forest only surviving towards the centre and west of Amazonia and also towards the east, with a large 'savanna corridor' between these two areas.
Sediments in the Amazon 'delta'; an indicator of conditions inland? Large amounts of arkosic (feldspar-rich) sand turn up in the Amazon submarine cone, deposited during glacial intervals. The presence of feldspar fragments in Amazon delta sediments was initially taken by Damouth & Fairbridge (1970) as indicating dry conditions during the glacial phase; however Irion (1984) has pointed out that the feldspars cannot have come from surface soils of the low-lying Amazon Basin, as required by the aridity hypothesis, because weathering from earlier humid periods should have chemically removed feldspars from the land surface before it became arid. Irion instead suggests that the feldspars were derived from the Carajas area and from river downcutting due to the fall in sea level. Damuth & Fairbridge (1970) also suggested that these could have been derived from the Guyana and Brazil Highlands, but it was pointed out by Milliman et al. (cited by Clapperton 1993) that these sediments might have been transported down from the Andes in the west of Amazonia (perhaps indicating aridity in the east Andean region?). At present, most of the Amazon sediment is derived from the Andes.
Furthermore Milliman et al. (cited and discussed in Clapperton 1993) suggested that the chemical maturity of many of the glacial-age sediments that are derived from coastal rivers (not the Amazon itself) indicates that strong chemical weathering on land had continued through glacial intervals, thus indicating that conditions remained humid during glacials. However, Clapperton (1993) points out that these coastal rivers carry very little sediment under present conditions, because the complete cover of rainforest prevents erosion. Thus, he suggests, an opening up or loss of forest cover (under drier conditions) would have been necessary to allow the stripping of the chemically weathered materials originally formed under humid interglacial conditions.
Colinvaux's view of a moist and cool glacial Amazon region. In contrast to the fairly generally held view of large-scale forest regression from the Amazon Basin during the LGM, Colinvaux and colleagues have instead advocated that the Amazon region remained predominantly moist. Colinvaux points out that the regions which show evidence for either a general climatic drying or specifically a loss of rainforest at around the last glacial period are either climatically marginal for rainforest nowadays (e.g. Carajas) or are of an altogether different and independent climate zone (e.g. Galapagos). At Mera, a site at raised elevation in the Napo region of Ecuador (near the eastern edge of Amazonia), Liu and Colinvaux (1987) have found pollen and wood evidence of a moist no-analogue forest of Andean and rainforest elements existing at 33,000 years ago and 26,000 years ago (radiocarbon dated), under conditions at least 4 °C cooler than today. Note that these dates have since been strongly disputed by Heine 1994, who finds evidence that the deposits are much older. At a site in southern Ecuador (Bush & Colinvaux 1989), in the foothills of the Condor Range, pollen and wood evidence dated to 31,000 years ago and 26,000 years ago present a very similar picture to the Mera site.
It may be relevant to note here that hiatuses are present in both of these Ecuador cores around 25,000-17,000 years ago. Whilst it is necessary to be very cautious in interpreting an absence of data, the lack of deposition might perhaps indicate some 'hidden' dry climatic event fitting into a pattern that other workers have suggested (pers. obsv.n., based on discussion with various people working on the region). A sedimentary hiatus occurs in many other places in South America at around the LGM raising the suspicion that a coherent climatic pattern of aridity was responsible, rather than simply local factors (M-P. Ledru, pers. comm. June 1993). In other parts of the tropics (e.g. Australia, Africa), a pattern of moist conditions around 30,000-25,000 years ago was often followed by aridity around 20,000-18,000 radiocarbon years ago. In SE Asia, montane ever wet sites do not seem to show the drying and retreat of forest that occurred in adjacent lowlands. On the other hand, if there was indeed a significant arid event in the western Amazon area at around the LGM, it must have ended very rapidly to allow moist forest to return to this site by 17,000 years ago when deposition resumes; this must put some doubt on the hypothesis of aridity in this case at least. However, it will be necessary to bear in mind the substantial doubt which has now been placed on the age of one of the two west Amazon sites by the dating reanalysis (mentioned above) carried out by Heine (1995).
Widespread hiatuses and terminations in cores around and within the lowlands of the Amazon Basin (rather than the raised elevation areas that Colinveaux's team have been sampling) for the LGM period may indicate rather more arid-than-present conditions in many places (pers. comm. M-P. Ledru ORSTOM Sept. 1992). However, the lack of deposits stretching back to the LGM may also be due to widespread river downcutting due to the lower sea level at that time, and also the high rate of reworking of floodplain sediments by the rivers (C. Clapperton pers. comm. 1991). The central Amazonian cores of Colinvaux's group show rainforest vegetation persisting through the LGM, but of course this only shows for certain that the central Amazon remained forested, which is really not a controversial issue (almost everyone working on South America agrees that there would have been rainforest in the central Amazon region at the LGM). What is more controversial nowadays is how far the surrounding rainforest of the Amazon region regressed.
Rather than emphasising aridity as an influence on Amazon forests at the LGM, Colinvaux and colleagues suggest that cooling was of great importance. On the basis of the cooling indicated by forest compositional changes at high and low altitudes in the Andes and central America, Colinvaux suggests that what we might define as 'modern-day' lowland rain forest was confined to below 500m or even 300m relative to present sea level (pers. comm. April 1990). Forest at higher altitudes than this would have been of a semi-temperate community of which there is no modern analogue (Colinvaux pers. comm. May. 1990).
It is difficult to gauge from his writings just how much rainforest/moist forest Colinvaux has in mind for the LGM Amazon. It is of interest to note, however, that Colinvaux's co-worker M.B. Bush has recently published a map suggesting for sake of argument that an overall 20% reduction in rainfall be considered for the LGM Amazon (Bush 1994). This would be enough to cause a significant retreat of rainforest, causing for instance a broad NW-SE trending savanna corridor to open up in eastern Amazonia. There would also be a significant loss of forest in SW Amazonia. The actual loss of forest area is not as great, however, as in other map reconstructions such as those published by Clapperton (1993) and van der Hammen and Absy (1994) (see below).
Recent research by Colinvaux's group is summarized in this linked web page. They report unpublished work from the upper Rio Negro (central Amazonia), which shows no sign of savanna encroachment during the last glacial (though some lowering of water levels). However, this is still within the area that most authors who advocate a large-scale forest retreat would suggest as remaining as forest during the LGM period. Note that many other authors suggest that the evidence of aridity and savanna encroachment in other areas of the Amazon Basin indicates a significant decrease in forest area during the last glacial (with gallery and floodplain forests dominating the pollen flora that turns up in the Amazon fan). This issue will hopefully be resolved more clearly over the next few years as more sites across the interior of Amazonia are located.
Van der Hammen and Absy's maps; a more drastic rainforest retreat. On the basis of the palynological and sedimentological evidence, Van der Hammen and Absy come to the conclusion that a more drastic reduction in rainfall of around 1000mm (a 40% decrease) would be necessary to eliminate forest from all three of the Carajas, Katira and Georgetown sites. Thus they suggest that if rainfall all across Amazonia fell by the same absolute amount it would leave scattered pockets of forest at the eastern coast and in southern Amazonia, and a large block of forest surviving in western Amazonia. Based on both palynological and sedimentological evidence of aridity, Clapperton (1993) also suggests in the form of a sketch map a large reduction in LGM Amazon forest cover, with a broad savanna corridor separating two main forest refugia in west and east Amazonia respectively. Clapperton's distribution of forest refugia is different from that suggested by Van der Hammen and Absy (1994), and may be based on extrapolation from different present-day rainfall maps (the actual mean rainfall distribution of rainfall across Amazonia is still subject to large uncertainties due to the sparsity of monitoring stations; P. Jones, University of East Anglia pers. comm.). The simpler east-west rainfall gradient assumed by Clapperton appears to be based on present-day rainfall maps similar to those given in recent editions of the Times World Atlas. His general LGM scenario of a central savanna corridor is followed here on the basis of my further consideration of the distribution of sites, but with greater forest cover to allow for the recent north-central Amazonian Rio Negro site which apparently remained forested through the LGM. Assuming that rainfall gradients were generally similar to the present distribution (in theTimes Atlas map) but at a different level, we have tentatively suggested LGM survival of rain forest in those areas which presently have above 1600mm annual rainfall.
The Mato Grosso. Desert conditions? In the present scrub/savanna region to the south of the Amazon forest, there are abundant signs of past aridity, although it is not clear whether this actually dates to the LGM. In the Patanal depression of the Mato Grosso Plateau, stabilised dunes occur even where the rainfall exceeds 1200mm, together with large alluvial fans and the remnants of clay or salt pans (indicating surface wash from a sparsely vegetated landscape, with periodic heavy storms such as occurs in semi-arid Australia at present). Thomas (1994 p.251) notes Klammer's statement that the landscape appears to be 'that of a desert come under the influence of a humid climate'. Klammer himself considered these forms to be pre-Quaternary, but Thomas (p.251) notes that Tricart and Ab'Saber have each suggested that they were late Pleistocene (LGM) in age. Better dating constraint on these deposits would be of considerable interest, to test whether or not they are of glacial age and more especially LGM age.
Semi-arid Eastern Brazil. Substantial cooling. Further east, in the present dry semi-arid tropical grassland/scrub zone of eastern Brazil (Piaui Province, 7 S, 41.5 W, 400m a.s.l.), noble gas levels in ancient groundwater suggest that temperatures at the LGM were 5.4 +/- 0.6°C cooler than during the late Holocene (Stute et al. 1995). This fits in with the general pattern from elsewhere across the American tropics.
Equatorial Andean region. Lowering of vegetation belts. A temperature depression of 6-8°C, at above 2000m at 5N, in Colombia is estimated from pollen data from Lake Sabana de Bogota (2550m a.s.l.), in the plain of Bogota (e.g. Van der Hammen & Gonzalez 1960, Hooghiemstra 1984). For the Eastern Cordiellera of Colombia, Hooghiemstra (1984) shows an LGM altitude profile based on extrapolating the temperate changes inferred from these high altitude pollen cores to the full range of altitudes (assuming the same lapse rate as today). In his reconstruction, tropical forest reaches up to 500m (presently 1000m), subandean forest is in a belt at 500-1300m (presently 1000-2500m), Andean forest at 1300-2000m (2500-3300m), paramo at 2000-3000m (3300-4000m), alpine desert at 3000-3500m (4000-4500m) and perennial snow above 3500m (presently above 4500m).
Based on a combination of palynological and geomorphological data, Heine (1995) suggests that in the Bolivian and Ecuadorian Andes, the temperature depression above 4,200m a.s.l. was 4.8°C relative the present. This temperature depression is slightly less than the estimates from the Colombian Andes, described above.
Drier than present on Inter Andean Plateau. Pollen data from sediments of a lake further to the south-west in the Ecuadorian Interandean Plateau (Colinvaux et al. 1988) shows drier as well as colder conditions than at present. The lowering the level of Lake Fuquene in the eastern Cordillera of Colombia suggests that rainfall at the LGM was at least 50% less than today (van der Hammen & Absy 1994). Colinvaux (1993) suggests on this basis that there was a modest increase in aridity in the adjacent Pacific coastal area as well.
But moister in the eastern cordillera of the Andes? Colinvaux (1993) notes in contrast, that the Bogota Lake referred to above shows a 'pluvial' pattern of higher water levels (moister climate) during glacials than during interglacials. He suggests that as it is on the eastern side of the Andes it is influenced by a separate climate system; one indicative of the moist LGM conditions that would have extended eastwards over lowland Amazonia. His pre-LGM submontane rainforest data from western Amazonia are taken as fitting into this general pattern. However, there appears to be a conflict with the pollen and lake level data present for the same general area by van der Hammen's group, above.
Subtropical South America.
Dry grassland/shrubs instead of humid forest. There is now some pollen evidence relating to the present coastal belt of transitional tropical rainforest and warm temperate forest of south-eastern Brazil. A lowland site at Poco Grande (26 deg.25'S, 48 Deg.52'W, 8m a.s.l.) going back to 14,000 14C y.a. shows substatially drier-than-present conditions with only rare rainforest elements (Alchornea, Urticales, Palmae) and an abundance of Myrtaceaeous trees (the latter possibly only reflecting localized swamp conditions, rather than any general forest cover) (Behling 1995). Conditions were perhaps even more arid previous to this, during the period including the LGM, judging by the possible timing of the aridity maximum shown for the adjacent uplands (see below). Behling notes that it is not yet possible to know whether a closed rain forest or merely scattered remnants persisted further north along the coastal plain.
In tropical SE Brazil, Behling & Lichte (in press) find pollen evidence of considerably cooler and drier conditions than present between about 48,000 and 17,000 14C y.a. (with a slight moistening of climate - indicated by slightly higher proportions of tree pollen - between 17,000 and 10,000 14C y.a.). This is at a site (20 deg.S, 43 deg.W)l, with the present vegetation being semi-deciduous forests and woodlands (present annual rainfall 1000-1500mm), replaced during the glacial period by grasslands with warm temperate trees existing only along rivers as gallery forests. A cooling of around 5-7 deg.C is suggested, with a movement of temperature zones at least 750 km equatorwards, and general replacement of forests in the region by grasslands throughout the glacial period (Behling & Lichte, in press).
Arid and cold in the adjacent uplands.Behling (1995) gives results of three cores from the uplands in SE Brazil, scattered through the area between about 26-29 deg.S and 49-50 Deg.W. When deposition started at around 14,000 14C y.a., grassland (campos) predominated, with Araucaria apparently very rare and confined to valleys. There are also various coarse slope wash deposits constrained with varying degrees of precision by radiocarbon dating to the period around the LGM, within the uplands between about 22-20 S and 45-50 W (summarised by Clapperton 1993, and Thomas 1994). Clapperton takes this evidence as indicating a sparsely vegetated landscape, probably arid grassland, subject to episodic heavy storms.
Dune fields in eastern Brazil? Thomas (1994 p.258) mentions Ab'Saber's recent work on dune fields in the Sao Francisco Valley (around 45 W, 15 S) of eastern Brazil (presently savanna vegetation), some 500 km south of the present north-eastern dry zone. Thomas mentions that the age of the dunes is uncertain, but that 'they appear young and probably relate to the LGM'. If this view is correct, the area would seem likely to have had a semi-desert or desert vegetation, with sparse ground cover.
Drier conditions in the subtropical woodland region. In what is presently the subtropical woodland region of south-central Brazil (19 S), the pollen-bearing LC3 core from Salitre (Ledru 1992) gives indications of vegetation up to 25,000 years ago and since 17,000 years ago, but with a hiatus in between. Ledru interprets the vegetation at around 17,000 years ago as existing under drier conditions than at present, with less forest vegetation than during the Holocene (high levels of arboreal pollen in her core are interpreted as being due to a localised expansion of Myrtaceae on the exposed lake bed, rather than a general expansion in forest cover), although at the time of most recent communication she felt that the data were not sufficient to allow her to know just how much forest cover there was generally in that region, because Salitre is the only dated core in this part of Brazil so it might not be representative of that region in general (Ledru pers. comm. 1993), although other more recently obtained cores from Behling (see above) confirm a general pattern of greater aridity. Nor is it possible to know what the LGM vegetation was like at this site. A decrease in sediment grain size towards the hiatus that occurs around the LGM is suggestive of decreased stream runoff, perhaps reflecting drier conditions (Ledru pers. comm. May 1994). This seems to fit in with the general pattern of more arid conditions across all or most of SE Brasil, which is suggested from the recently obtained cores of Behling and colleagues (see above) in the forest regions nearer to the coast.
Colinvaux (pers. comm. June 1993) mentions the unpublished 1992 thesis of De Oliveira, with a continuous record from the south-eastern cerrados area, at Minas Geras, stretching back to well before the LGM. This area now has scrubby cerrados vegetation, but during most of the glacial Colinvaux (pers. comm. 1993) reports that the climate was apparently moister and cooler. Although Colinvaux does not specify in his letter whether the LGM itself was moister and cooler, or just the phase preceding it, the implication is that conditions at this site were indeed relatively moist at the LGM. Note however that Clapperton summarises an array of geomorphological evidence of sand dunes, dune fields and blowouts from the southern Cerrados region, and suggests extensive sand desert at the LGM for the area bordered by the Beni Basin in the north and the Pantanal in the east, and the northern Cerrados region in the south.
Retreat of Araucaria forest in SE Brazil. Another site from the SE Brazilian highlands within the present Araucaria forest zone, at Morro de Itapeva (22 deg.47'S, 45 deg.32'W), 1850 m a.s.l., suggests markedly cooler and drier than present conditions up until about 17,000 14C y.a. (Behling in press). High-elevation grasslands predominated, and Arauacaria forest elements were rare. Conditions apparently ameliorated somewhat after 17,000 y.a.
Drier vegetation in the sub-equatorial Andes. In the central Peruvian Andes at 11 S, palynological evidence from Laguna Junin shows that the present Andean forests were poorly represented there and that sub-puna shrublands took their place, presumably due to cooler and drier conditions than at present (Villagran 1990, Markgraf 1993).
Replacement of forest by savanna. In the extreme north-east of Argentina (Misiones Province), pollen evidence indicates that instead of the sub-tropical rainforest vegetation that now covers the hills there was a savanna vegetation (Iriondo & Garcia 1993; citing a 1985 reference in Spanish by Rizzo). Whether this was relatively dry or moist savanna is not specified. If this relatively high rainfall area had suffered such a reduction in rainfall, surrounding areas that are drier than this must also have been affected, having correspondingly drier vegetation than occurs today.
Stevaux (1994) reports the preliminary results of palynological studies of the Upper Parana River, Porto Rico site at the southern end of the broadleaf subtropical forest zone (22 deg.S, 53 deg.W), and compares these results with those of other pollen sites nearby. During the late Glacial the zone was apparently dominated by savannas and grasslands instead of forest. Geomorphological evidence also indicates aridity; there was a braided river system and colluviation. Stevaux suggests savanna vegetation possibly persisting until around 6,000 y.a.
Loess deposition occurred across Santa Fe and South-western Entre Rios, which Irondo & Garcia suggest as indicating much drier conditions than at present. Iriondo & Garcia suggest a north-easterly movement of the present climate isoclines by about 750 km under LGM conditions (see below). This would correspondingly roughly to a northward shift of the zones by 6 Latitude, and bring temperate semi-desert conditions over most of northern Argentina.
Desert and semi-desert in Chaco. Further to the west, in the western Chaco plain (northern Argentina and Paraguay) that lies next to the Andes, the LGM climate was dry enough to allow mobilisation and then deposition of aeolian sand and loess. Clapperton (1993) reviews the evidence of dunes and blowouts in this area, and also the area to the north of the Chaco (in southern Bolivia, W. Paraguay and N. Argentina), where he reconstructs sand desert for the LGM. On adjacent areas to the south-east, a thick loess built up during the last glacial, covering the whole plain from the flank of the Andes as far as 60W. Animal fossil evidence suggests a dry steppe or semi-desert environment in this loess belt (Iriondo & Garcia 1993). Estimates of discharges from the palaeochannels of westward flowing the rivers in this plains region indicate flows that were only about 20% of present levels (Iriondo & Garcia 1993).
Patagonia and the south of the Andes.
Frigid, dry conditions at the southern tip. In the high southern latitudes of South America, ice sheets or ice caps were present along the Andes, spreading down onto the plains and coasts below. The surrounding vegetation in this most southerly part of the peninsula (at around 45 - 51S) seems to have been a windy dry heath-like steppe-tundra, existing under a cold climate with low precipitation (Heusser 1987, V. Markgraf pers. comm. 1993).
Replacement of Andean mediterranean zone by steppe woodland. Further north, west of the Andes at Laguna Tagua Tagua (at about 34 S), steppe vegetation (with some evergreen Nothofagus trees present) existed over at least part of the area now occupied by mediterranean-type sclerophyll woodland (Heusser 1983). There is no sign of closed forest persisting in the area; the vegetation at this locality seems to have been an 'open evergreen forest' according to Markgraf (1989) or 'steppe woodland' as Heusser describes it, with similarities to the open woody vegetation that occurs near the forest limit in the mountains today. In a sketch map drawn for us by C. Heusser (pers. comm, 1990), a park tundra grading northward into steppe-woodland is shown between about 40 and 30S.
Atacama desert persisting? Meanwhile, what survived of the Chilean mediterranean vegetation had moved further equatorwards. The most northerly extension of the Chilean mediterranean vegetation during the last glacial seems to have been about 30 S, with localised presence of sclerophyll taxa in coastal ravines at that time (Villagran 1990). Further to the north of this, the continuing presence of the Pacific coastal/montane Atacama Desert through both warm and cold periods of the Quaternary is supported by present-biogeographical evidence; the physiognomically similar altiplanic and mediterranean floras of northern and central Chile remain floristically very distinct, suggesting that the aridity barrier between them has remained unbroken for millions of years (Villagran 1990). Clapperton (1993) suggests that the desert belt would also have extended further north along the coast that at present, in line with the indications of generally more arid conditions elsewhere along the Pacific edge of South America.
The Chilean temperate rainforest zone; did it disappear? Further south along by the Pacific coast (approximately in the zone depicted by the Heussers as park tundra at the LGM), at Isla Chiloe (42S), conditions in what are presently the evergreen temperate rainforest belt also seem to have been wetter than now, with replacement by the hyperhumid magellanic moorland down to sea level (Villagran 1988). It thus seems that some aspects of the vegetation zones presently in this area were shifted by at least 5 latitude equatorwards at the LGM, with the moorland vegetation descending in altitude at least 600m relative to its present lower limit. However, it seems, according to the Heussers, that the temperate evergreen rainforest (in its various forms) did not shift correspondingly northwards in this way, but was lost as a vegetation unit. Localised forest tree refugia were probably present, in the form of woodland with steppe (L. Heusser pers. comm. 1990) between about 35 and 48S. Markgraf (1991) instead presents a map in which the area 40-35S that the Heussers depict as 'park tundra' and 'steppe-woodland' is described as 'Nothofagus dombeyi' woodland. Note that there is no mention of any areas of surviving 'forest' vegetation in Markgraf's reconstruction. As Markgraf's (1991) work is the most recent, I have here based the map reconstruction here mainly on her published maps.
Note however that the reconstruction of Villagran (1990) disagrees with this view of a complete loss of the west Andean temperate evergreen forests; her diagrams show large areas of evergreen forests persisting further north than their present ranges. This is based on her interpretation of increased rainfall further north due to a shift in the westerly winds (see below), on the basis of the same pollen evidence (summarized by Markgraf 1993) that the Heussers and Markgraf have been citing.
Was precipitation in southern Chile greater at the LGM? There is some disagreement over the extent to which precipitation and/or moisture availability might have increased relative to the present at the LGM in some areas of southern Chile. Heusser (1983) and Villagran (1989) advocate a precipitation increase caused by a northwards shift of the rain-bearing wind belt. However, Markgraf (1989) is more in favour of relatively greater LGM aridity in many areas and no significant increase in precipitation in other areas. She suggests that cooling of climate promoted the growth of ice caps and favoured moister conditions in some areas of the lowlands simply due to decreased evaporation. However, according to Villagran (1989) there may have been a very substantial increase in precipitation (at least 1500mm) over the area where magellanic moorland formed at the LGM. She suggests a temperature decrease of about 4°C relative to the present, whilst Markgraf (1989) suggests a cooling of 6°C for the general area of southern Chile.
Eastern side of the Andes; drier than present conditions in Patagonia, the pampas and adjacent areas. In an extensive review of the palaeoevidence of this region, Iriondo and Garcia (1993) suggest a very substantial drying and cooling over this whole region, stretching down into Patagonia. Precipitation and temperature isoclines seem to have been some 750 km to the north-east of their present positions.
In central Argentina the mean annual temperature may have been around 2.5-3°C lower, with the difference in winter (4°C) being greater than in summer (1°C), according to Iriondo & Garcia (1993). They report that rainfall generally seems to have been about half of what it is today (although it is not clear whether the temperature decrease has been allowed for in this calculation). In contrast, Thomas (1994 p.217) cites the evidence of Gonzales that after 17,500 years ago the level of Lake Salina del Bebeder (33S, Argentina) was high. However, Thomas does not mention whether or not the lake level indicates aridity actually at the LGM itself.
Iriondo & Garcia suggest in their climate-zone maps that the north-east of Argentina remained 'subhumid' through the LGM, although they do not specify what sort of vegetation would have been associated with this.
Desert in the pampa. In what are presently the 'pampa' dry grassland areas of western Argentina, periglacial silts and sands washed down from the Andes were blown away to form a sand sea covering the south-western half of the Pampa, indicating that there may have been almost no vegetation (Iriondo & Garcia 1993). The LGM sand sea extended between 38 and 33 deg.S (Latrubesse & Ramonell 1994) and included large longitudinal dunes more than 100 km long (Ramonell et al. 1992).
To the east, large areas of loess acumulated north to 30 deg.S. This loess was deposited in a semi-desert environment, (derived from the desert areas in the west) and covered over the pre-existing drainage system of the plains (Latrubesse & Ramonell 1994).
Absence of forests on the eastern edge of the Andes. On the eastern (Argentine) side of the temperate evergreen zone of the Andes at 37-42S, areas that are now covered with high or low elevation evergreen beech forests were dominated before 12,000 years ago by non-arboreal pollen and only minor amounts of xeric trees and shrubs, presumably due to aridity (Markgraf 1989). Markgraf suggests, on the basis of this palaeovegetation pattern observed from west to east across the Andes, that the precipitation gradient was even greater than today's.
Northward movement of Patagonian vegetation. To the north-east, in what is presently the semi-desert scrub zone of Argentina (south of the Chacos), at a site 100km east of the Andes (34S), a Patagonian semi-desert flora of herbs and grasses seems to have been present, associated with a cool, dry climate (Markgraf 1989).
8,000 years ago (early Holocene).
Central America.
Forests in place. In Central America, forest lines had risen to their present altitudes before 8,000 years ago (Hooghiemstra et al. 1992). At Chalco Lake in the high altitude plateau of central Mexico, temperate montane forest had replaced the earlier grasslands before 9,000 y.a. (Lozano-Garcia et al. 1993). In the Peten region of Guatemala, conditions became moister before the start of the Holocene but the expansion of tropical forest elements seems to have been accompanied by persistence of some temperate tree taxa until about 9,000 years ago. By 8,000 years ago the present tropical semi-deciduous forest had fully returned to the Peten region of Guatemala, and persisted throughout the Holocene (Leyden et al. 1993). Colinvaux (pers. comm, Aug. 1993) suggests on the basis of his pollen cores that there may have been drier-than-present conditions in lowland Panama during the early Holocene.
Significant human disturbance? Piperno et al. (1990) point to an abrupt increase in charcoal fluxes and pollen of 'weedy' species in Lake Yeguda, Panama, in the early Holocene (11,000 years ago) as indicating the onset of deliberate disturbance by either cultivators or hunter gatherers. Colinvaux (1993) suggests that this may indicate a significant role of human disturbance in the forests throughout this period. However, agricultural impact may have been very limited. Maize pollen does not begin to appear in pollen diagrams before about 6,000 years ago.
Northern coast of South America. Moister than present in Venezuelan highlands. In cores from Lake Valencia in the Venezuelan highlands, moist tropical forest was present in the earlier Holocene, whilst the area today has a markedly more seasonal climate (Salgado-Labourieu 1977, Colinvaux 1987).
Amazonia.
Forest in west-central Amazonia. Lake cores from the western-central Amazon show that the forest there has been in place since at least 7,000 years ago, which is as far back as the cores go (Colinvaux pers. comm.), which at least does not contradict the view that it would already have returned by 8,000 years ago. The slow rise in the nitrate level in the Andean Huascaren Glacier core (Thompson et al. 1995) suggests that Amazonian forest (a major source of nitrate, as opposed to savanna which is not) did not in fact reach its maximum extent until the mid-Holocene, although nitrate levels were already much higher than during the last Glacial by early Holocene time. The view of a delay in forest colonisation fits in with some of the palynological evidence reviewed below.
Forested, but drier in eastern Amazonia. In the eastern Amazon area, the core CSS2 from the plateau of Southern Serra dos Carajas (Absy et al. 1991), shows drier-than-present conditions around 8,000 years ago, correlative with a reduction in Amazon River discharge. An increase in savanna elements occurs at this time. This early-to-mid Holocene dryness is also observed in other parts of South America; in Brazil (Servant et al. 1989) and in the high Andean lake Titicaca, in Bolivia (Wirrmann et al. 1988).
However, the overall forest cover might not have been much different from the present-potential; a recently obtained lake core (Sifeddine et al. 1994, manuscript submitted) also on the Serra dos Carajas plateau in the rainforest region of the eastern Amazon (Brazil) shows that at 8,000 years ago the area was about as forested as at present, with a mainly evergreen rainforest cover. The forest here did not reach this climax state until around 9,500 years ago (strongly lagging the lake level rise, which began around 13,000 years ago, and showing an abundance of early successional species for several thousand years). Behling (1995) reports that during the lower Holocene and late Glacial, an 'unknown ecosystem' rich in Ilex with some Podocarpus occurred in the east Amazon coastal area, though he does not state until what time during the Holocene this no-analogue community persisted. He also reports human disturbance (charcoal layers) from closed forest at the Amazon mouth starting at around 10,000 years ago.
Forest line in Andes. The work of van der Hammen suggests that the highest forest limit in the equatorial Andes was reached after 8,000 years ago (indicating a maximum in temperature perhaps 1 or 2°C higher than today), with some drop towards modern times.
Moister in Atacama Altiplano? The upland coastal high desert of Peru and Chile, (extending between 18 and 28 S at present) seems to have been greatly reduced by higher rainfall at some stage during the late glacial or early Holocene. Precipitation at present is well under 200mm, but according to loosely dated glacial and lake sediments it would have exceeded 500mm, reducing the desert to a zone between 25 and 28 S (Grosjean & Messerli 1995). However, they also find indications that by the mid-Holocene, conditions were even more arid than at present.
The South American subtropics.
Moister than today's climates, after a late switch from aridity. In Beunos Aries province of eastern Argentina (about 35S) and in the Cordoba uplands at about 30S in the west of Argentina, the climate seems only to have switched from glacial aridity to Holocene moistness at around 8,500 years ago, according to sedimentological, archaeological and zoological evidence cited by Iriondo & Garcia (1993). The same wide range of sources of evidence suggest conditions that were strikingly moister than today's at 8,000 years ago, and throughout the mid Holocene.
A subtropical forest expansion?
In south-central Brazil, semi-deciduous forest predominated in the area of the Salitre core of Ledru (1992). The presence of Araucaria forest up until 8,000 years ago suggests the absence of a dry season (Vernat et al. 1994). Around 8,000 years ago, winter temperature (from ostracods & diatoms) seems to have been about 7°C, or 2-3°C cooler than today, and moister with only a short dry season, with the conifer Araucaria as a dominant component in the forest cover.
But less forest in other areas. Drier-than-present conditions - with less forest than during the late Holocene - may have existed in the subtropical forest area of SE Brazil from after about 8,000 years ago until about 3,000 years ago, indicated by charcoal phases followed by savanna incursion (Vernet et al. 1994). Other evidence of more arid than present conditions at the 8,000 y.a. timeslice comes from a pollen record at Lago do Pires; 17 deg.57'S, 42 deg. 13'W (Behling 1995b). A herb savanna seems to have predominated, with gallery forests in the valleys, in contrast with the latest Holocene when closed semideciduous forest covered the whole area. Thus, the maps reconstruction I have given here, with expanded early and mid Holocene forest in SE Brazil, may be incorrect.
But much more humid further south? Iriondo & Garcia suggest that from 8,000 years ago to 3,500 years ago the humid subtropical forest climate that is now present only in the extreme north-east of Argentina would have covered much of north-eastern Argentina, from just north of Buenos Aries eastwards, across Uruguay. Thus in the north-east, precipitation seems to have been around double what it is today (Iriondo & Garcia 1993), which would give totals of more than 1600mm.
Note however that there are indications of various dry periods, with slope wash deposits, during the Holocene (see discussion by Thomas 1994, p.254-255). The timing and duration of these episodes is unknown.
Woodland/forest in the Argentinean central region? Likewise, subtropical humid conditions extended south-westwards; near the coast at Cipoletti, 39S and 60W, annual precipitation seems to have been about five times its present amount (at that time having a 7-month wet season), with a mean temperature about 1.8°C warmer than today's (the greatest difference being in the winter temperature) (Iriondo & Garcia 1993). This would presumably have been associated with widespread forest or woodland cover (my reconstruction) in this central and northern belt of Argentina, which is presently semi-arid and lacking a dense vegetation cover. As an example, at the southern edge of this extended zone, a lake pollen core at 35S 58W showed the existence of a humid steppe with rainforest elements 8,500-4,000 years ago; presumably, forest cover would have been increased north-eastwards of this site. Over the same period, an essentially Brazilian subtropical mammal fauna extended south to 38 'and probably further' (Iriondo & Garcia 1993).
Moist steppe in the west. In the western part of Argentina, north of Patagonia (about 38S) and in the 200-300km wide belt east of the Andes (north to about 25S) , there seem to have been humid enough climates for widespread formation of 'black earth' soils that survive where they were buried by later Holocene windblown sediments (and in some places as relict surface soils). These soils were presumably originally associated with a rich steppe vegetation. This is in areas that have presently have only a sparse vegetation cover and little or no soil development (Iriondo & Garcia 1993).
Patagonia staying arid. Markgraf considers that the area south of 50S stayed in an arid mode, but with a woodland vegetation, until after 9,000 years ago (Markgraf 1992), and perhaps as late as 8,000 years ago (Markgraf 1992). At latitudes between 50 and 55 S, the postglacial forest expansion seems to have started at around 9,000 years ago in the west, and to have reached over to the east side of the Andes by 8,000 years ago (Markgraf 1992). After this, moister conditions, slightly warmer than today's (with a continuing spread of Nothofagus beech forest) set in gradually during the early-to-mid Holocene (Heusser 1990). However, for the extreme southern tip of South America, essentially modern vegetation distributions from around 13,000 years ago onwards have been reported (e.g. Villagran 1990).
5,000 14C years ago.
Amazonia. Arid episodes and wildfire. Colinvaux (1987) suggests the possibility of a dry mid-Holocene phase of rainforest regression and replacement by savanna, occurring intermittently during the interval 7,000-4,000 14C years ago. Various short-lived episodes of aridity and of wildfire have been documented in pollen diagrams (Carajas and Matto Grosso) and charcoal layers (Venezuelan Amazon) have been found for the mid-to-late Holocene, apparently antedating the arrival of Amerindians in these particular sub-regions. A recently published study also suggests similar drought and fire episodes for central Amazonia (near Manaus) between 5,000 to 7,000 years ago (Piperno & Becker 1996), without any indicators that these were caused by cultivation. However, there does not seem to be evidence that any single arid phase lasting for more than 50 years, and most seem to have been much shorter (Colinvaux 1987), though Thomas & Thorp (1992) report that Markgraf has found lake level evidence for an arid phase lasting several hundred years at around 6,000 years ago in tropical America. Suguio et al. (1993) discuss the evidence for dry phases in the 7,000-4,000 year ago interval on the basis of the Serra dos Carajas core at 6 S, 50 E in Eastern Amazonia. At this site, rainforest regression seems to have occurred in the form of frequent dry episodes (marked by charcoal fragments) alternating with times of partial forest recovery with high percentages (>40%) of the pioneer vegetation genus Piper. At the Carajas site, savanna does not seem to have had any time to encroach between times of forest pioneer influx, suggesting that the drought episodes were severe but short-lived.
Central America.
Agriculture not yet important as a disturbance factor? At 5,000 years ago (and as early as 7,000 years ago), agriculture seems to have existed in the Central American region but does not seem to have been an important influence on the vegetation. In the forested Peten lowlands of Guatemala, Binford et al. (1987) find initial increases in erosion and decreases in tree pollen only after around 3,000 years ago. Markgraf (1993) suggests that widespread agricultural disturbance in Central America occurred only after about 3,500 years ago. Similarly, Piperno (1994) finds that for various sites in lowland Panama, forest disturbance by humans was very minor and sporadic up until around 4,000 years ago, after which major forest loss began to occur. In a contrasting environment, in the uplands of central Mexico (Basin of Mexico), large scale deforestation by humans apparently occurred only after 3,000 y.a. (Lozano-Garcia et al. 1993).
Sub-tropical South America.
Mire development commencing. Heusser (1990) finds relatively cool, mesic conditions similar to the present beginning with the development of beech forest and mires starting around 5,000 years ago, in a core in southern Tierra del Fuego (55S).
Warmer and moister than today in some areas, but drier in others?
Iriondo and Garcia (1992) find evidence of conditions still having been moister than at present, at 5,000 years ago being much as they were at 8,000 years ago, with a large north-eastward displacement relative to the present vegetation.
However conditions might have been drier than today, further to the north. For the subtropical forest area of south-eastern Brazil at Salitre (Suguio et al. 1993), there was a minimum in tree pollen (to around 10% of the total; the present tree pollen % for this woodland site is around 15%) at around 5,000 years ago, indicating a longer dry season than at present. The presence of charcoal and savanna grasses further reinforce the view that conditions were substantially drier than at present. Hence on further consideration, the map reconstruction here showing an expanded area of subtropical forest is likely to be incorrect. Using various bio-indicators, winter mean temperature is estimated to have been around 5°C higher than the present value of 10°C (Suguio et al. 1993).
There is an apparent contrast between the pattern of seasonally drier conditions to the north, and the indications of moister-than-present conditions in Argentina. Thus, Iriondo & Garcia (1993) suggest that in Argentina and adjacent areas, conditions at 5,000 years ago were much moister and slightly warmer than today, with the vegetation-climate boundaries similar to those at 8,000 years ago (see above).
Drier than present in the Chilean Andes. At subtropical latitudes, three high-altitude lakes in Northern Chile (respectively 18 S, 27 S & 23 S) show a pattern of mid-Holocene dryness, based on isotope studies of ostracodes (Schwalb & Burns 1995). The dry episode is suggested to be between about 7,000 and 4,000 years ago, perhaps linked to persistent 'El Nino' events. The spatial extent of the aridity is not known, and it contrasts with the indications of moister conditions in Argentina (see above).
Names of QEN participating experts (named in the text above) who have made direct contributions to this work on South America:
Hermann Behling, Center For Marine Ecology, Fahrenheitstrasse 1, 28359 Bremen - Germany, Tel: x49 421 2208 321, Fax: x49 421 2208 330, E-Mail: hbehling@zmt.uni-bremen.de
M.B. Bush, Department of Zoology, Ohio State University, 1735 Neil Avenue, Columbus, Ohio 43210, USA. (E-mail: MBUSH@OHSTVMA ).
C. Clapperton, Department of Geography, University of Aberdeen, Aberdeen AB9 2UF, UK.
P. Colinvaux, Department of Zoology, Ohio State University, 1735 Neil Avenue, Columbus, Ohio 43210, USA, and Smithsonian Tropical Research Institute, UNIT 0948, APO AA 34002-0948, USA.
T. van der Hammen, c/o Tropenbos Colombia, Sede Corporation Aracuara, Apartado Aereo 34174, Bogota, Colombia.
C.J. Heusser, New York University, New York, USA.
L. Heusser, New York University, New York, USA.
H. Hooghiemstra, Hugo de Vries Laboratory, University of Amsterdam, Kruislaam 318, 1098 SM, Netherlands.
M. Iriondo, CONICET, Parana, Argentina.
M-P. Ledru, ORSTOM, Centre de Bondy, 72 Rte d'Aulnay, 93143 Bondy cedex, France.
S. Lozano-Garcia, Instituto de Geologia, National University of Mexico, 04510, Mexico D.F., Mexico.
V. Markgraf, University of Colorado, Boulder, Colorado, USA. e-mail; markgraf@spot.colorado.edu
A. Rancy, e-mail; Alceu@cfh.ufsc.br
M. Sarnthein, Christian-Albrechts University, Germany.
B. Turcq, ORSTOM, Centre de Bondy, 72 Rte d'Aulnay, 93143 Bondy cedex, France.
{kind=link}
{kind=link}